![]() Go to frame view (Recommended only for
screen resolution 1024x768)
Go to frame view (Recommended only for
screen resolution 1024x768)
8.1.6 Supercoiling of DNA
In the early sixties, J. Vinograd and coworkers found that the DNA of some bacteriophages and mitochondria may exist in the form of cyclic molecules. As was established later, most viral and many cell DNAs have a circular form. When both strands in a circular molecule formed by a duplex DNA are covalently closed (a similar situation arises when the ends of loops formed by a duplex DNA are linked by proteins), they are topologically constrained and can no longer be separated.
The most important parameter of each DNA molecule covalently closed into a circular structure is the linking number. It is denoted Lk0 (or
a) and equals, to a first approximation, the number of intersections of the two strands in a double-stranded ring (Fig. 8-11). Thus, Lk0 has integer values. The most important point is that Lk0 is invariant for a given covalently closed circular DNA.If such a DNA is in a relaxed state - that is, it can lie in one plane without any constrain - then Lk0 = Tw, where the winding number Tw (or
b0) equals the number of turns in the double helix of the DNA.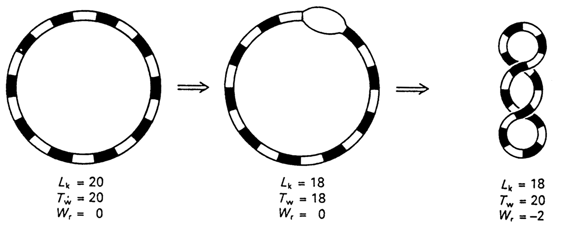
Fig. 8-11. Introduction of two supercoils into DNA with twenty duplex turns (adapted from C. R. Cantor and P. R. Schimmel, Biophysical Chemistry, part Ill, W H. Freeman and Co., San Francisco, 1980).
However, if the number of turns in the double helix is increased or decreased without affecting the covalent continuity of both strands, then, by virtue of the constant nature of Lk0, such a change will be compensated by formation of supercoils (Fig. 8-11). The number of supercoils in a superhelical DNA is denoted Wr (from "writhe") (or
t) and can be written asWr can be a positive or a negative quantity. Covalently closed circular DNAs isolated from cells are negatively supercoiled as a rule. When Wr is negative, the linking number of the duplex DNA is lower than the number of double helix turns in a linear DNA of the same length or, in other words, the two strands in a negatively supercoiled DNA are "underwound" with respect to each other. The degree of supercoiling of a particular DNA can be best characterized by the density of supercoils (also called "specific linkage"),
s, which equals Wr/Tw. For most naturally occurring supercoiled DNAs s is in the neighborhood of -0.05. It should be borne in mind, however, that in reality we always deal with populations of DNA molecules with different Lk0. Therefore, s is the mean density of supercoils for a given DNA preparation.Supercoiled DNA can be observed by electron microscopy (Fig. 8-12). The number of supercoils in DNA is usually determined by gel electrophoresis. The separation is based on the fact that the greater the value of Wr, the more compact the DNA macromolecule and, consequently, the greater the electrophoretic mobility in the gel (see, for example, Fig. 8-14 in section 8.1.7).
From the functional standpoint it is important that supercoiled DNA is higher in free energy than the relaxed form. Hence, local unwinding of a double helix of DNA with negative supercoils will lead to a release of the stress due to supercoiling and, therefore, it is energetically advantageous. Its clear manifestation can be seen in the fact that negative supercoiling strongly stimulates the transition from the right-handed B form of DNA into left-handed Z-DNA. Indeed, already under normal physiological conditions segments with sequences d(CG)n . d(CG)n and d(AC)n . d(GT)n incorporated into DNA with
s = -0.06 are transformed into a left-handed Z form.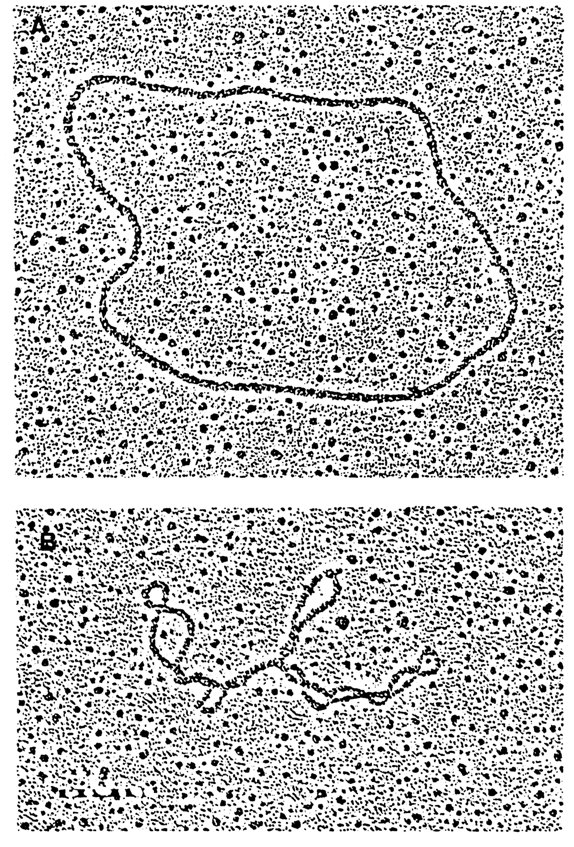
Fig. 8-12. Electron micrographs of (A) relaxed and (B) supercoiled circular DNAs (courtesy of G. G. Gause).
In the cell, the negative supercoiling of DNA is provided by DNA gyrases which belong to the category of topoisomerases. Topoisomerases are enzymes capable of altering the number of supercoils in covalently closed circular DNAs. They are subdivided into two groups: topoisomerases I and II. The substrate for topoisomerase 1 is a supercoiled DNA in which they reduce the number of supercoils and eventually turn it into relaxed circular DNA. Topoisomerases 11 (DNA gyrases), on the contrary, can create DNA supercoils.
When this occurs, every single step of DNA gyrase action results in two negative supercoils and its activity is associated with hydrolysis of ATP. Topoisomerases are present in every living cell and play a vital role in the processes of DNA replication and recombination.
8.1.7 Unusual DNA Structures
Another example of the influence exerted by supercoiling on the structural changes in a double helix of DNA is provided by formation of cruciform structures. Virtually every DNA contains inverted, or palindromic, repeating sequences varying in length from several to many thousands of base pairs.
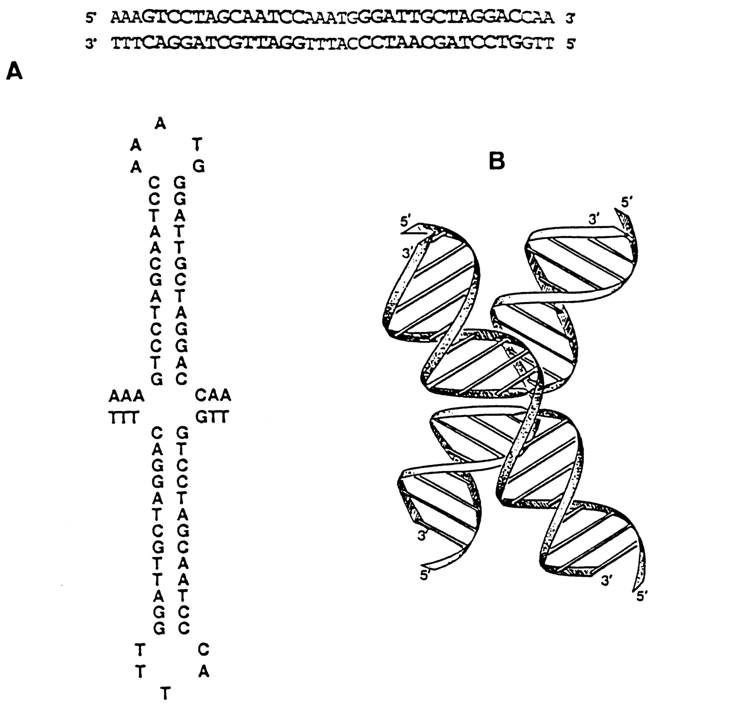
Fig. 8-13. (A) Linear and cruciform structures of the inverted repeat sequence of Col E1 DNA. (B) 3D model of the cruciform structure (adapted from A. 1. H. Murchie and D. M. J. Lilley, Methods in Enzymol., 211, 158-180 (1992)).
Theoretically, one can picture transformation of a linear duplex palindrome to a cross (Fig. 8-13). In the case of a relaxed DNA, the probability of such a transformation is negligible. Since it is energetically advantageous in DNA with negative supercoils, cruciforms in vitro have been observed in all examined supercoiled DNAs with a normal density of supercoils. (Experimentally, cruciform structures are identified as having single-stranded loops in the apices of "hairpins" which are cleaved by nucleases specific with respect to single-stranded DNA.) The formation of cruciforms can also be demonstrated by gel electrophoresis (Fig. 8-14). The question as to occurrence of cruciform DNA structures in vivo remains open. The rate of their formation is extremely low, which is possibly why nobody has been able to observe it in the cell.
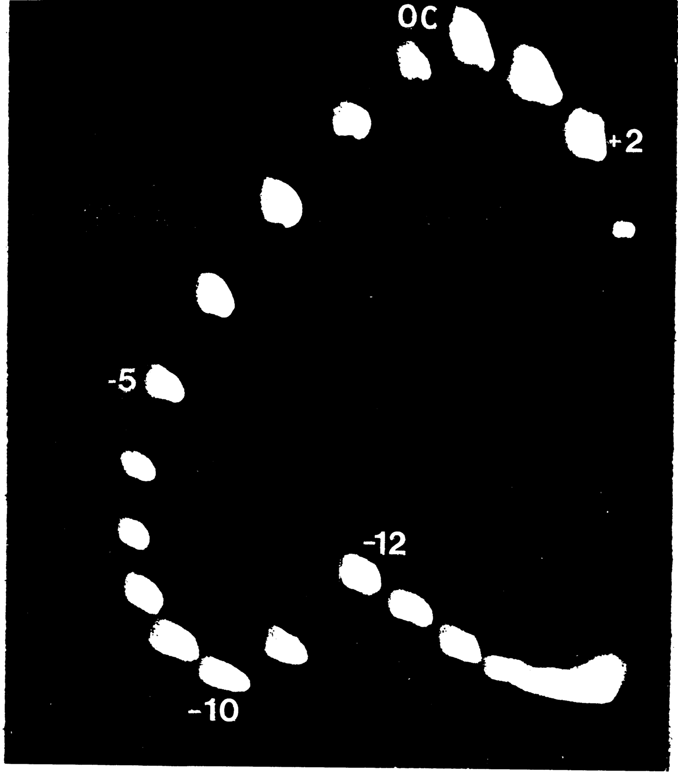
Fig. 8-14. Separation of DNA topoisomers by two-dimensional gel electrophoresis. After separation in the first direction (from top to bottom along the left edge), the gel was saturated with an intercalator (see 8.1.8) which reduced the number of supercoils in the DNA. Then, electrophoresis was conducted in the second direction (from left to right). The upper spot (OC) corresponds to relaxed DNA. Positively supercoiled topoisomers move clockwise; negatively supercoiled topoisomers move counterclockwise. The difference in mobility between topoisomers -10 and -12 means that topoisomers -12, -13, and so on have a cruciform structure (courtesy of M. D. Frank-Kamenetskii).
If DNA contains homopyrimidine-homopurine sequences, negative supercoiling may turn it into a form illustrated in Figure 8-15. Since in the case of sequences of the d(AG)n . d(CT)n type such a transition occurred at low pH values (and at pH 4.3 it is observed even at
s = 0), it was termed the H form. The H form has been given as the reason for existence of segments in natural DNAs overly sensitive with respect to nucleases specific toward single-stranded polynucleotides.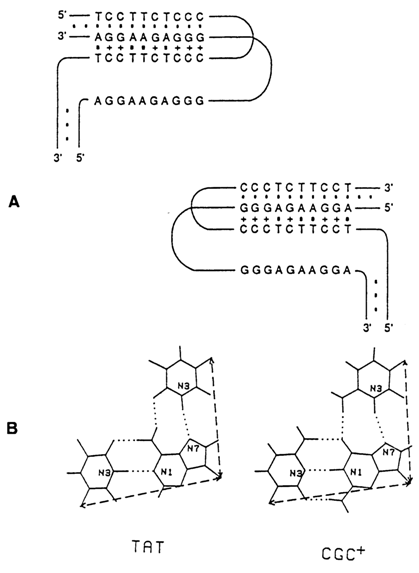
Fig. 8-15. (A) Two isoforms of H-DNA. (B) Scheme of base triads (adapted from M. D. Frank-Kamenetskii, Methods in Enzymol., 211, 180-191 (1992)).
Unusual structures of DNA which do not require supercoiling to arise by virtue of the ability of guanine bases to form tetrades. They are responsible for the structural organization of telomers - special segments created at the ends of linear chromosomes by the telomerase enzymes. Telomers consist of repeating sequences which contain blocks of three or four guanines. G-rich sequences associate with one another to form four-stranded structures known as G-DNA. Among the many possible G-DNA structures we selected as an example the antiparallel dimers shown in Figure 8-16, which contain guanosines in both syn- and anti-conformation. It can be seen that at first singlestranded G-rich segments fold back and then the two hairpin loops associate into G-DNA.

Fig. 8-16. Structure of G-DNA formed with (dG), stretches within single-stranded DNA segments (adapted from J. R. Williamson, Curr. Opinion in Stuct. Biol., 3, 357-362 (1993)).
Unusual DNA structures were studied using a great variety of techniques. Chapter 9 will present some evidence of formation of such DNAs through chemical modification.
8.1.8 Interaction of Ligands with Double Helices of DNA
DNA forms complexes with proteins both in vivo and in vitro which affect its function. Moreover, numerous low-molecular weight substances, primarily antibiotics and carcinogens, interact with DNA and materially affect its functional properties. Many of such substances are mutagens. With few exceptions, proteins as well as low-molecular weight ligands are bound to DNA in a noncovalent fashion.
The ligands interacting with DNA can be divided into two groups according to the manner in which they are bound to the double helix - groove binders and intercalators. The first group includes a great variety of proteins, primarily those regulating transcription processes. Some low-molecular weight substances containing positively charged groups are also bound to grooves of the double helix. A highly distinctive feature of the second group of ligands is their having polycyclic aromatic groups with various side chains.
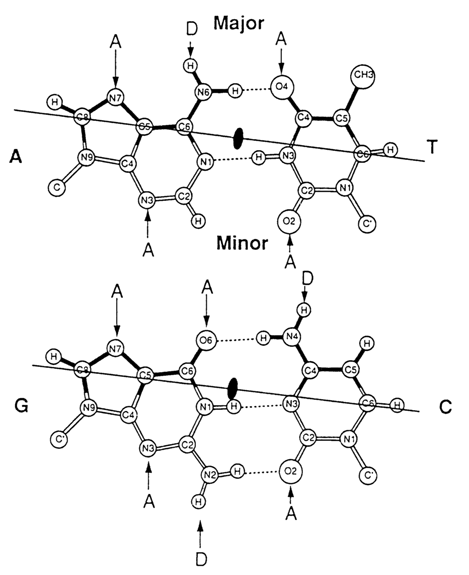
Fig. 8-17. Donors (D) and acceptors (A) of hydrogen bonds in grooves of the double helix of DNA (adapted from T. A. Steitz, Quart. Rev. Biophys, 23, 205-280, D (1990)).
In view of the great functional importance of DNA-ligand interactions it would be appropriate here to discuss their mechanism in greater detail.
Groove Binders. A remarkable feature of this group of ligands is their ability to recognize nucleotide sequences in DNA without unwinding its double helix. This recognition is possible due to formation of specific hydrogen bonds between the functional groups of the ligands and those of the heterocyclic bases "looking" into grooves of the double helix.
Analysis of Figure 8-17 which illustrates Watson-Crick pairs easily brings to light a marked difference between the patterns of proton-donor and -acceptor sites in the minor and major grooves of the double helix of B-DNA. It can be seen that the protein in the major groove (or any other groove binder) is capable of readily recognizing any of the four base pairs (A . T, T . A, G . C and C . G). On the other hand, the ligand in the minor groove can discriminate base pairs only by composition (i. e., G- and C-containing ones from those containing A and T). Therefore, the recognition of the nucleotide sequence in DNA by regulatory proteins takes place in the major groove of the double helix.
Figure 8-18 offers an example of specific contacts formed by the amino acids of a regulatory protein with the heterocyclic bases in the major groove of DNA. The protein shown here regulates the transcription process and features what is known as helix-turn-helix motif. One of the et helices of this feature fits into the major groove of DNA. Note that the proteins also interact with the minor groove, which renders the complexes highly stable.
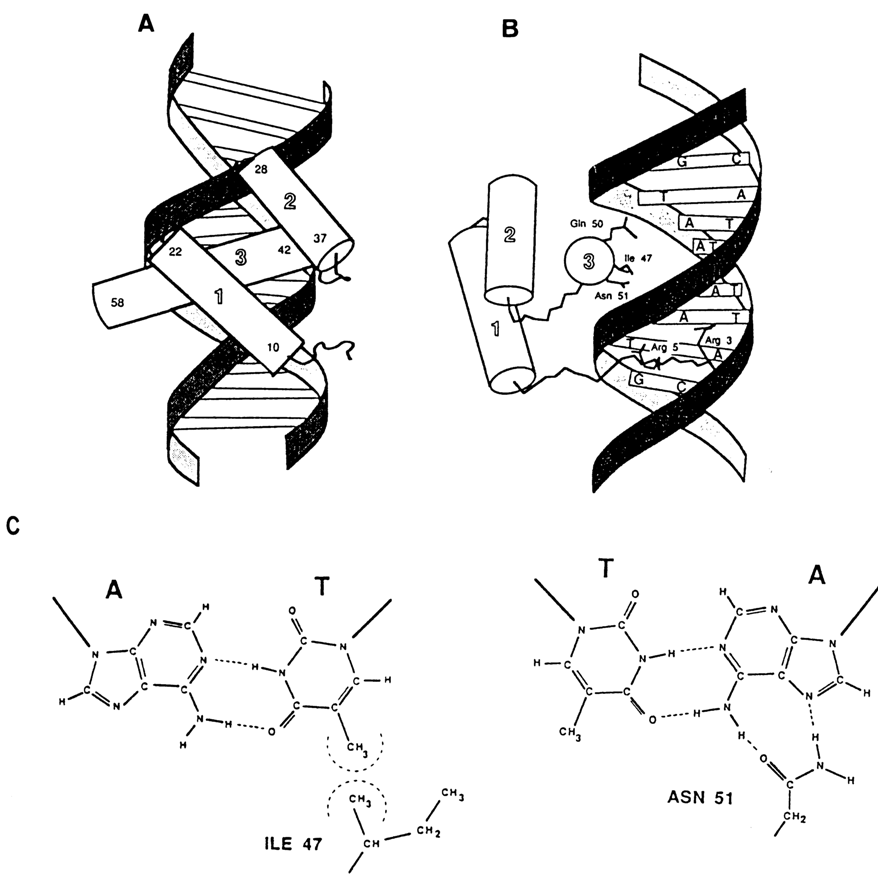
Fig. 8-18. DNA-protein complexes with a helix-turn-helix motif of so-called homeodomain in a Drosophila protein that regulates development. (A) and (B) - two projections of the complex showing the arrangement of helices (shown as cylinders) and the N-terminal arm with respect to the double-helical DNA; C - formation of hydrogen bonds between T . A pair and Asn-51 at hydrophobic contact between A . T pair and Ile-51 respectively (adapted from C. K. Kissinger et al., Cell, 63, 579-590, (1990)).
Another example of binding between a ligand and the minor groove of the double helix is provided by the complex of DNA with the antibiotic netropsin (Fig. 8-19). The peptide groups of the antibiotic form hydrogen bonds with the 2-keto group of thymine and N3 of adenine. Therefore, the antibiotic becomes specifically bound to A-T-rich regions of DNA. The netropsin DNA complex is also stabilized by electrostatic interactions between the positively charged guanidinium groups and negatively charged phosphate groups of DNA.
It would be wrong to assume that groove binders do not affect at all the initial conformation of the double helix of DNA. As the double helix of DNA interacts with regulatory proteins, it exhibits a rather important ability to bend (bendability) and even form kinks, which enhances its contacts with the protein molecule. Moreover, groove binders bring about local changes in the mutual arrangement of the bases in complementary pairs, and that results in, among other things, local changes in groove size. Since, as we have already pointed out, B-DNA displays microheterogeneity dependent on the nucleotide sequence, deformations of the double helix are sequence-dependent, too.
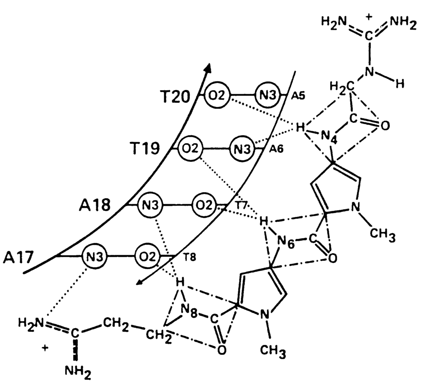
Fig. 8-19. Scheme of specific interactions between netropsin and A . T pairs in the minor groove of the double helix of DNA (adapted from M. L. Kopka et al., J. Mol. Biol., 183, 553, (1985)).
Intercalators. Intercalation implies incorporation of a polycyclic
aromatic compound between two base pairs into DNA without affecting not only the covalent
linkages in their polynucleotide chains but also the Watson-Crick pairing. As far back as
1961, L. Lerman proposed a model according to which intercalation of a planar polycyclic
dye into the double helix of DNA increases the distance between neighboring base pairs
from 3.4 to 6.8 Å (Fig. 8-20). The model suggested that intercalation must be accompanied
by a change in the angle of rotation of base pairs about the helix axis, which is to say
that the helix is unwound. Besides, intercalation is, of course, accompanied by elongation
of the helix. Later, the model was fully borne out by direct X-ray structural analysis of
single crystals of complexes formed by oligonucleotide duplexes with various
intercalators. One such complex is illustrated in Figure 8-21.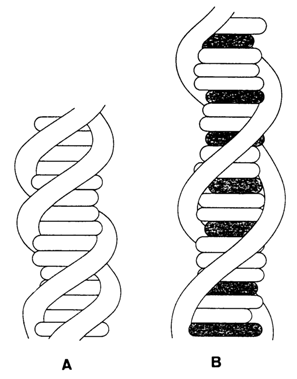
Fig. 8-20. Lerman's intercalation model: (A) native DNA; (B) complex of DNA with a polycyclic aromatic dye (black disks).
In the case of covalently linked cyclic DNA molecules, intercalation alters the number of superhelices in their supercoiled forms. For instance, intercalation of the dye ethidium bromide, widely used for this purpose, into the double helix of each DNA molecule unwinds it by 260. As can be inferred from the relation between Wr, Lk0 and Tw (8.1.6), binding of ethidium bromide to a natural supercoiled DNA (for which, as is known, Wr < 0) will first lead to disappearance of the superhelices and emergence of a relaxed form, followed by coiling of the double helix in the opposite direction. Indeed, according to the results of sedimentation analysis (Fig. 8-22), as the intercalator concentration increases, the supercoiled DNA molecule becomes less and then more compact.
Some intercalators exhibit rather high selectivity of binding to DNA. For example, the antibiotic actinomycin D widely used as a transcription inhibitor binds exclusively to C . G sites of B-DNA. If an interealator contains side substituents, they occupy the grooves of the double helix to form, as a rule, specific contacts with heterocyclic bases and, thereby, make the interaction between the intercalator and DNA even more specific. A similar complex is formed by DNA with the antibiotic daunomycin which contains a sugar moiety.
Finally, the ability to be incorporated into the double helix of DNA may be displayed by an intercalator together with the capacity to form covalent bonds with DNA. If an intercalator is a bifunctional cross-linking agent, it forms cross-links between the DNA strands. The most widely used for this purpose are both natural and synthetic psoralens - linear bifunctional tricyclic furocoumarines which photoreact with DNA to form cyclobutane adducts with pyrirnidine bases.
Fig. 8-21. Two projections of a ethidium-dinucleotide complex, drawn on the basis of X-ray analysis; (A) looking along the mean planes of the base pairs; (B) looking down on these planes (adapted from S. C. Jain and H. M. Sobell, J. Biomol. Structure and Dynamics, 1, 1161-1194 (1984)).
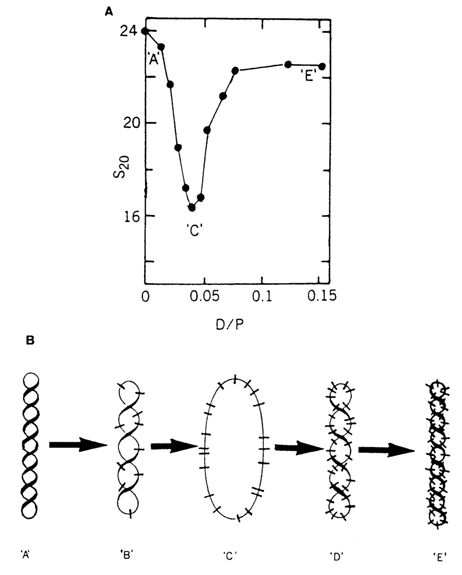
Fig. 8-22. (A) Titration of supercoiled DNA with ethidium bromide; (B) Interpretation of the esults presented in (A). D/p = dye bound/nucleotide. 'A'-supercoiled DNA; 'B'-partially unwound DNA; 'C'-relaxed DNA; D'-partially overwound DNA; 'E'-overwound DNA (adapted from W Guschelbauer, in: Encyclopedia of Polymer Science and Engineering. 12, 699-785 (1988)).