![]() Go to frame view (Recommended only for
screen resolution 1024x768)
Go to frame view (Recommended only for
screen resolution 1024x768)
7.5 Complexes of Polynucleotides with Mono- and Oligonucleotides
Single-stranded polynucleotides may form ordered helical complexes with complementary nucleosides, mono- and oligonucleotides. Their study is important for elucidating the mechanism of the interaction of nucleotide derivatives and short segments of nucleic acids with polynucleotide templates in the course of protein and nucleic acid synthesis. Moreover, as will be shown below, complexes of the oligomer-polymer type are widely used to investigate the secondary and tertiary structure of nucleic acids.
The formation of complementary monomer-polymer complexes can be observed only in the case of purine nucleosides and nucleotides. Pyrimidine nucleosides and nucleotides do not form such complexes. This suggests that stacking interactions (which are more pronounced in the case of purine derivatives) are essential for stabilization of the complex.
Analysis of the stoichiometry of monomer-polymer complexes indicates that under optimal conditions triplet complexes are usually formed, which are more stable than the corresponding double-stranded complexes. In other words, in the case of an adenosine-poly(U) system, for example, a complex is formed made up of two poly(U) strands and a sequence of adenosines (it can be regarded as poly(A) without phosphodiester bonds).
Naturally, in the case of mononucleotides (i.e., in the presence of charged phosphate groups), monomer-polymer complexes may form only in solutions with high ionic strength (when the phosphate groups are screened).
Oligonucleotides form much more stable complexes with the complementary polynucleotide, as compared to nucleosides and mononucleotides. Therefore, formation of oligomer-polymer complexes is observable not only when purine oligonucleotides but also pyrimidine ones are involved.
Oligomer-polymer complexes exhibit all the properties inherent in the polymer-polymer structures discussed in the previous section. The most interesting property is the dependence of the complex stability on the oligomer length. Here, the role of stacking interactions in complex stabilization becomes especially manifest. Indeed, as has been established for an oligo(A) . poly(U) mixture, all oligomer-polymer complexes beginning with the trinucleotide are characterized by hypochromism amounting to 35 %. A similar hypochromic effect is observed when a triple-stranded poly(A) . 2poly(U) complex is formed. Thus, it may be assumed that in the oligo(A). poly(U) complex oligomers form a continuous chain with the terminal links interconnected through stacking interactions.
On the other hand, in an oligo(U) [or oligo(dT)]-poly(A) system the oligomer-polymer complex displays the same percentage hypochromism as the poly(U) . poly(A) duplex, only beginning with the 14- to 16-unit oligonucleotides. It is believed that in the oligo(U) part of the complex two or three bases at each end are not involved in stacking, which precludes formation of a "continuous" helical complex.
Incorporation into the oligomer of bases noncomplementary with respect to those of the polymer disturb the ordered structure of a complex and destabilizes it.
Such complexes have been of particular interest because their formation underlies the site-specific mutagenesis technique. Experiments with short oligodeoxyribonucleotide complexes have shown that the behavior of an unpaired nucleotide is strongly dependent on the heterocyclic base species. For instance, in the duplex
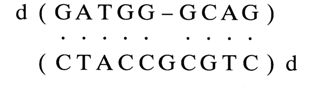
the "extra" guanosine is intercalated into the double helix and participates in the stacking interactions with the adjacent GC-pairs. In this case, the helix is slightly bent at the site of the defect. Similar behavior is displayed by adenosines, for example, in the duplex
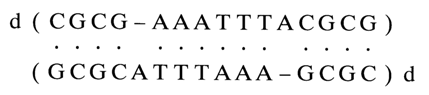
when it is in solution. In the crystalline state, the duplex is marked by protrusion of the unpaired adenosines from the double helix. This is indicative of a certain conformational flexibility of double-helix polynucleotide complexes with unpaired bases.
An unpaired cytidine may also exist two states - outside the helix and incorporated into it, the transition from the former to the latter state occurring when the solution is heated. Unpaired uridines always tend to be outside the double helix. In contrast, thymidine, which is more likely to be involved in stacking interactions, is incorporated into the helix without causing any perceptible changes in its conformation.
It is known that complementary base pairs at the termini of duplexes are in a partially denatured state or, to be more precise, in equilibrium between the paired and unpaired states. Such termini are said to "breathe". If a helix terminates in several unpaired bases (as is the case, for example, with DNA fragments derived with the aid of restriction endonucleases), the stability of the double helix is adversely affected. Interestingly, however, single unpaired terminal nucleotides enhance the stability of the duplex. It has been speculated that this is due to additional stacking interactions between the unpaired and paired bases in the terminal complementary pair. For example, the melting point of the duplex
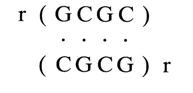
in a buffer containing 1 M NaCl is 27.50 C, whereas in duplexes
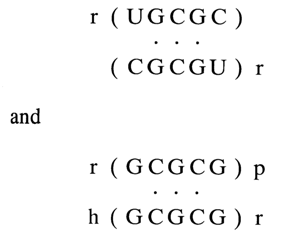
in the same buffer it is 31.5 and 510 C, respectively. Consequently, the increase in melting point of the duplex is indeed in direct correlation with the ability of the unpaired base to participate in stacking interactions.