![]() Go to frame view (Recommended only for
screen resolution 1024x768)
Go to frame view (Recommended only for
screen resolution 1024x768)
7 Conformation of Nucleic Acid Components. Macromolecular Structure of Polynucleotides
The current concepts concerning the structure of DNA and RNA provide the basis for research into the mechanisms of gene expression. In turn, elaboration of three-dimensional DNA and RNA structure models, their proof and further refinement are inexorably associated with advances in the structural chemistry of nucleic acids. Moreover, the fact that Watson and Crick had come up with the double helix model of DNA was testimony to the spectacular progress achieved in this branch of the chemistry of natural compounds in the early fifties.
It is absolutely clear that three-dimensional models of the structure of synthetic and natural polynucleotides can be constructed and correctly interpreted only with a profound knowledge of the conformation of monomer components of nucleic acids - nucleosides and nucleotides.
The conformation of nucleosides, nucleotides, oligo- and polynucleotides is studied using a wide range of physical and physico-chemical methods of which the most prominent are X-ray structural analysis and NMR spectroscopy. Also quite useful in these studies have been such simple spectral techniques as ultraviolet spectroscopy, optical rotatory dispersion (ORD) and circular dichroism (CD) as well as theoretical conformation analyses.
This chapter deals primarily with the final results produced by these approaches, without dwelling on the underlying theoretical concepts. Therefore, more detailed information on the latter can be obtained in the excellent textbook by C. Cantor and P. Schimmell (see references at the end of the chapter), which contains everything one needs to know about the theoretical fundamentals of these methods and their application to the structure of nucleic acids and their components.
7.1 Conformation of Nucleosides and Nucleotides
7.1.1 Heterocyclic Bases
All of the five major bases forming part of nucleic acids are more or less planar. As can be seen from the example illustrated in Figure 7-1, the atoms of heterocyclic rings are less than 0.01 nm (0.1 A) away from the mean ring plane.
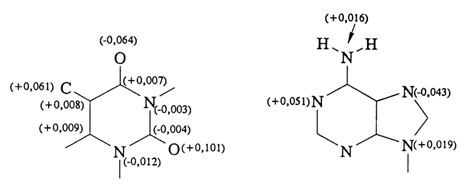
Fig. 7-1. Maximum displacement of atoms from a mean plane in thymines (deoxythymidine 5'-phosphate) and adenine (deoxyadenosine).
The planar conformations of heterocyclic bases are quite consistent with their chemical properties - all constituent bases of nucleic acids are quasiaromatic compounds.
A remarkable exception from this rule is dihydrouracil which lacks aromatic properties and has a half-chair conformation, the
C5 and C6 atoms being spaced 0.03 nm (0.3 A) apart (Fig. 7-2).According to X-ray structural data, in most nucleosides and nucleotides the C1' atoms of the carbohydrate (ribose or deoxyribose) moiety, involved in glycosidic bonding, are 0.01 to 0.02 nm (0.1-0.2 A) away from the mean plane of the heterocyclic ring.
The exocyclic amino groups of purine bases and cytosine are involved in the formation of resonance structures. and the C-NH bond has properties making it close to a double bond. Consequently, rotation about this bond is hindered, and the exocyclic amino groups together with the associated heterocycles are coplanar (lie in the same plane). At the same time, the exocyclic C=O group displays normal double-bond properties.
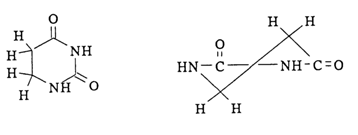
Fig. 7-2. Puckered conformation of the dihydrouracil ring.
7.1.2 Ribose and Deoxyribose
As regards pentafuranoses, which form part of monomer units in nucleic acids, it has been established that four atoms of the five-membered ring are more or less coplanar in most cases, whereas the fifth atom, C2', or C3', is 0.05 to 0.06 nm (0.5-0.6 A) away from the ring plane. In this case, if the C2', or C3' atoms lie on the same side of the ring plane as C5' we are dealing with so-called C2'- and C3'-endo-conformations, respectively; and if C2', or C3' are not on the same side as C5' we have C2'- and C3'-exo-conformations.
Such conformations in the ribose and deoxyribose moieties of nucleosides and nucleotides become clear if we address ourselves to such classical objects of stereochemistry as cyclopentane and tetrahydrofuran.
It is known that the planar form of cyclopentane is energetically unfavorable. Its molecule has conformations with one or two adjacent carbons away from the ring plane (Fig. 7-3).
In the case of tetrahydrofuran, whose derivatives are sugar moieties in nucleosides and nucleotides, similar non-planar conformations must occur as well. Calculations have shown that the most energetically favorable conformations for the pentafuranose ring are such in which the heterocyclic atom of oxygen and the neighboring C1' and C4' atoms are coplanar, while C2' and C3', do not lie in the ring plane. These conformations are shown in Figure 7-4.
An interesting exception is adenosine 3',5'-cyclic phosphate which has a C4'-exo-conformation.
It should be pointed out that in the literature the C2'-endo-conformations of carbohydrate moieties in nucleic acids are often referred to as S-conformations, while the C3'-endo-conformations are referred to as N-conformations. The reason is that the values of the pseudorotational phase angle lie in the range of 0 to 30' for the C3'-endo-conformations and 150 to 180' for the C2' endo-conformations or, in other words, in the northern (upper) and southern (lower) regions, respectively, of the pseudorotational cycle.
The majority of crystalline forms of nucleosides and nucleotides examined by X-ray structural analysis have either a C2'-endo or C3'-endo-conformation of their carbohydrate moiety. These conformations were identified in approximately equal numbers of cases, which suggested that the energy barrier
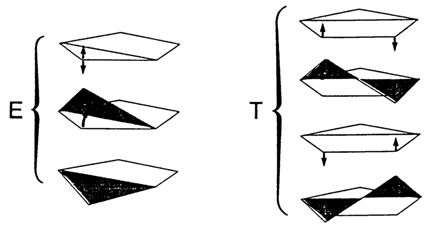
Fig. 7-3. Envelope (E) and twist (T) conformations of cyclopentane (adapted from W Saenger, Principles of Nucleic Acid Structure, Springer Verlag, N.Y, 1984).
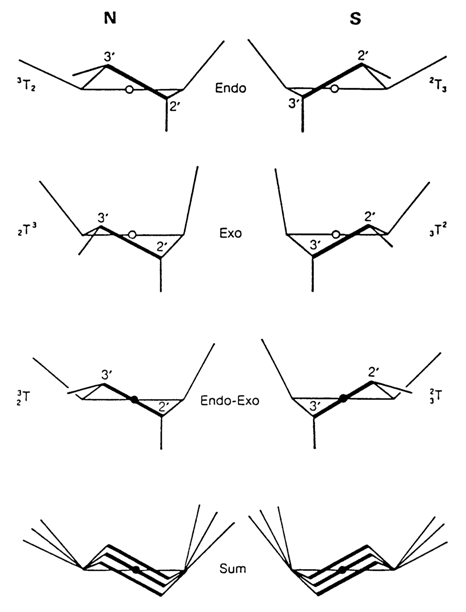
Fig. 7-4. Major puckered conformations of the pentafuranose ring of ribose and deoxyribose in nucleosides (adapted from W Saenger, Principles of Nucleic Acid Structure, Springer Verlag, N.Y, 1984).
between the two is low. Theoretical calculations indicate that it is only 5-10 kJ/mol (1-2 kcal/mol) for deoxynucleosides. In the case of ribonucleosides, this barrier must be perceptibly higher: 25-30 kJ/mol (6-7 kcal/mol), to be precise. Experimental NMR data attest to a rapid equilibrium between the N and S conformers in solutions of nucleosides and nucleotides.
The readily observed ("primary") deformation (puckering) of the furanose ring usually goes hand in hand with another, slighter ("secondary") one. For instance, in cytidine 3'-phosphate whose C2' atom lies nearly 0.05 nm (0.5 A) above the ring plane, C3' is 0.015 nm (0.15 A) away from the latter in the opposite direction. Thus, the ribose moiety virtually has a conformation which is intermediate between the C2'-endo- and C2'-endo-C3'-exo-conformations. An important consequence of deformation of the furanose ring is departure from the true cis-conformation of the 2'- and 3'-hydroxyl groups in ribonucleosides. The torsion angle between the bonds linking C2', O2' and C3', O3' is 43 to 540, whereas for true cis-bonds it must be equal to zero (see also Fig. 7.6.A for a definition of torsion angles).
X-ray structural analysis reveals three types of conformation for the C4'-C5' bond, which are illustrated in Figure 7-5.
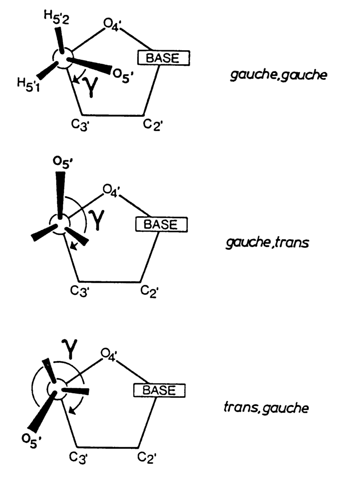
Fig. 7-5. Three possible conformations of ribose or deoxyribose due to different orientations of substituents about the C4'-C5' exocyclic bond (adapted from W Saenger, Principles of NucleicAcid Structure, Springer Verlag, N.Y, 1984).
In ribo- as well as deoxyribonucleosides, all three conformations are found. At the same time, the presence of a phosphate group at C5', in nucleoside 5'-phosphates hinders rotation about the C4'-C5' bond. As a result, all nucleoside 5'-phosphates are in a gauche-gauche conformation characterized by minimal interaction between the substituent at C5' and the furanose ring atoms.
As regards the conformation of the phosphate group itself, in nucleoside 5'-phosphates the phosphorus atom is usually in a transposition with respect to C4'.
7.1.3 Orientation of the Heterocyclic Bases Relative to the Sugar
An essential feature of the three-dimensional structure of nucleosides and nucleotides is the relative orientation of the base and sugar moiety, which is usually described by two parameters. The first parameter is the dihedral angle between the base and sugar planes (in this case, the mean square plane of the furanose ring is implied). In all nucleosides and nucleotides, it approaches, although never reaches, the right angle (ranging from 550 for adenosine 3'-phosphate to 810 for 5-iododeoxyuridine). The other major parameter is the torsion angle
c, or the angle of rotation of the base about the glycosidic bond. In addition to the angle c and puckered conformation of the sugar the threedimensional structure of the nucleotide in the polynucleotide chain is also given by six torsion angles of the main chain (Fig. 7-6B).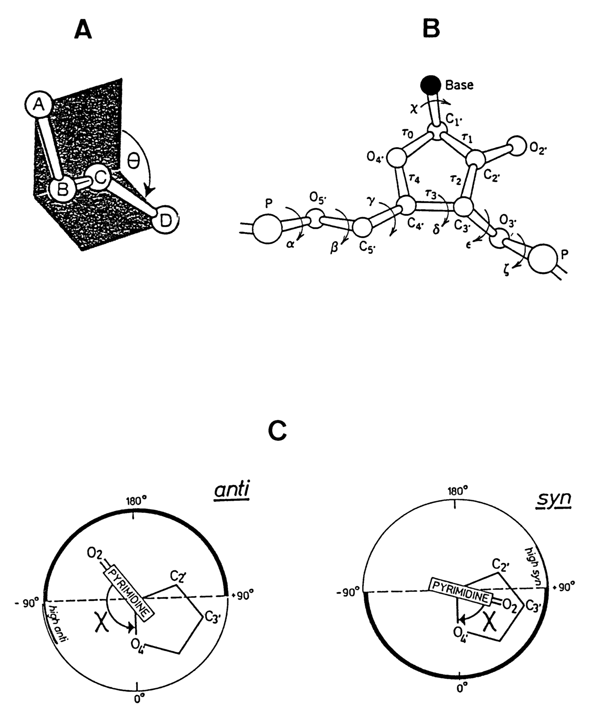
Fig. 7-6. System of torsion angles and their definition for a nucleotide in the polynucleotide chain. A - Definition of torsion angle
q (A-B-C-D) describing the orientation of bonds A-B and C-D relative to the central bond B-C. It can be seen that this angle is determined as that between planes A-B-C and B-C-D; B - Torsion angle c and nomenclature of six torsion angles along the phosphate-sugar-phosphate chain (courtesy of John Wiley & Sons, Inc.); C - Determination of the magnitude and sign of angle c for pyrimidine nucleotides in syn- and anti-conformations. The pyrimidine base is oriented toward the reader (consequently, atom C1' and bond N1-C2 are invisible) and rotated with respect to the immobile sugar (adapted from IUPAC-IUB Joint Commission on Biochemical Nomenclature, Abbreviations and Symbols for the Description of Conformations of Polynucleotide Chains, Eur. J. Biochem., 131, 9-15, 1983).Figure 7-6A shows how the torsion angles are determined. According to the rules adopted by the IUPAC-IUB in 1983, the torsion angle formed by the O4'-C1'-N1-C2 bonds in pyrimidines and O4'-C1'-N9-C4 bonds in purines equals 00 when the O4'-C1 bonds in the carbohydrate moiety lie in the same plane with the N1-C2 and N9-C1 bonds, respectively (Fig. 7-6C). It should be emphasized that one can find other (older) rules for determining the torsion angle in the literature.
The possible conformations of nucleosides, arising as a result of rotation of the base about the glycosidic bond, were first analyzed on molecular models using the so-called method of rigid spheres. This method resides in calculation of the distances between certain atoms in the base and carbohydrate moiety as functions of the angle
c.As was established using this approach, the strongest hindraces to rotation about the glycosidic bond in derivatives of the pyrimidine series occur as a result of interaction between the oxygen at C2 and hydrogen at C6 with the substituents at C2' and C3', on the one hand, and the endocyclic oxygen of the sugar, on the other. In the purine series, hindrances are caused by interaction between N3 and the same atoms of the furanose ring. As a general rule, however, the rotation of the base in purine nucleosides is hindered to a much lesser extent than in pyrimidine ones.
Careful conformation analysis of pyrimidine and purine nucleosides and nucleotides by this method has revealed two regions marked by the weakest intramolecular interactions. The first region corresponds to the angle equal to about -1000 and the other, +1000 (see also Fig 7-7C). The conformations associated with the first region are usually called anti-conformations and those associated with the second region are known as syn-conformations.
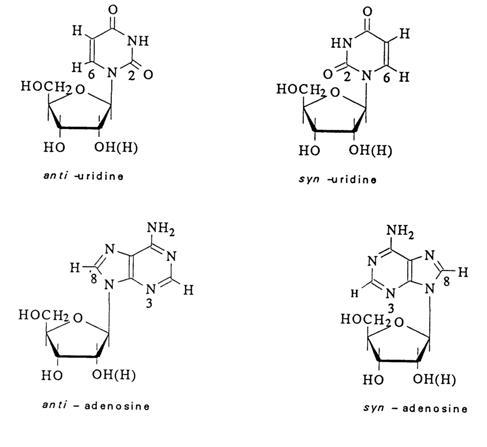
Fig. 7-7. Syn- and anti-conformations of nucleosides.
In other words, in the case of an anti-conformation in pyrimidine nucleosides it is the hydrogen at C6 that is the closest to C5' whereas in the case of an syn-conformation the closest atom to C5' is the oxygen at C2 (Fig. 7-7A).
In purine nucleosides, anti-conformation is marked by proximity of the hydrogen at C8 to C5', and in the case of syn-conformation N3 is above the plane of the carbohydrate moiety (Fig. 7-7B).
Later, theoretical conclusions were subjected to rigorous experimental verification by different methods. As indicated by X-ray structural analysis, crystalline pyrimidine nucleosides and nucleotides exist in anti-conformation in most cases. A classical exception from this rule is 4-thiouridine which crystallizes in the usual syn-conformation. It should be borne in mind, however, that when in solution, this nucleoside has an anti-conformation. Anti-conformation has also been found to occur in many purine nucleotides and nucleosides in the crystalline state. Guanyl ribo- and deoxyribonucleosides and -nucleotides present a special case because they are marked by syn-conformation both in the crystalline state and in solution. This, for example, is precisely what makes pdG and pG in oligonucleotides different from the rest of nucleotides. In this case, syn-conformation seems to be assured by the interaction between the 2-amino group of guanine and the 5'-phosphate group. Generally an anti-conformation can be converted in to a stable syn-conformation by inserting a large substituent at position 6 of the pyrimidine ring or position 8 of the purine ring.
The relative orientation of the base and sugar in nucleotides and nucleosides in solution was investigated in the early days primarily by optical activity measurements and, more recently, by NMR spectroscopy. In the former case, the object of the investigation was the optical rotatory dispersion or circular dichroism of nucleoside solutions in the UV region where these compounds are characterized by the Cotton effect. Since heterocyclic bases are optically inactive and ribose (or deoxyribose) does not absorb UV light in the region of 230 to 350 nm, it is believed that the sign and magnitude of the Cotton effect in nucleosides and nucleotides are determined chiefly by the relative orientation of the base and pentose. For example, pyrimidine nucleotides and nucleosides are characterized by positive Cotton effects near the absorption maximum of the pyrimidine base. A positive Cotton effect is a direct consequence of an anti-conformation in pyrimidine derivatives, since the cyclic derivative of uracil with a fixed anti-conformation also exhibits a positive Cotton effect.
Purine nucleotides display Cotton effects of smaller magnitude, as compared to pyrimidine ones. This finding confirms that the rotation about the glycosidic bond is easier in pyrimidine, rather than purine, derivatives.
More definite information on syn- and anti-conformations of monomer units in nucleic acids in solution can be obtained by NMR. This method makes it possible to easily identify NMR spectral peaks corresponding to protons associated with particular atoms of heterocyclic rings. As has already been mentioned, depending on whether a compound is in a syn- or anti-conformation, different protons of the heterocyclic base will be close to C5, (and the phosphate group in the case of nucleoside 5'-phosphates). Accordingly, the relationship between chemical shifts and pD (the NMR spectra were taken in heavy water) was studied for certain protons of the base. The results obtained for adenosine 5'-phosphate and its monomethyl ester are shown in Figure 7-8.
It can be clearly seen that at pD values exceeding 6.0, when secondary dissociation of the phosphate group begins, the value of the chemical shift changes only for the hydrogen atom linked with C8 (just as expected, no changes take place in methyladenosine 5'-phosphate). A similar effect can be observed only if adenosine 5'-monophosphate is in an anti-conformation.
Similar measurements were carried out for other purine and pyrimidine nucleoside 5'-phosphates as well, and in each case these compounds were found to be in anti-conformation.
There is a certain correlation between the syn- and anti-conformations of nucleosides and nucleotides, on the one hand, and conformation of their sugars, on the other. For instance, purine nucleosides with the C3'-endo-conformation of the sugar are usually in a syn-form. In the case of C2'-endo-conformation of the sugar, anti- and syn-conformers occur with equal probability. As regards pyrimidine nucleosides and nucleotides which, as has already been pointed out, are usually in anti-conformation, the angle
c depends on the conformation of the sugar as follows: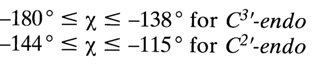
Comparison of the conformations of monomer units in nucleic acids in the series: "nucleoside"
® "nucleotide" ® "nucleotide of the polynucleotide chain" indicates that their conformational rigidity increases. This has prompted Sundaralingam to formulate the "rigid nucleotide" concept according to which nucleotides in nucleic acids belong as a rule to one of only two classes of conformers:
shown in Figure 7-9.
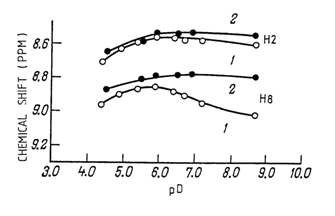
Fig. 7.8. Chemical shift 6 of different protons as a function of pD for adenosine 5'-phosphate (1) and its monomethyl ester (2).
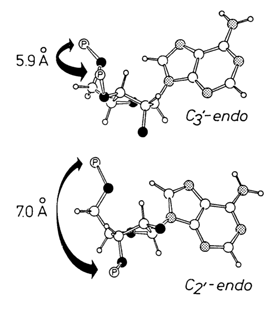
Fig. 7-9. Two conformations of a nucleotide in a polynucleotide chain. Note that in the case of the 2'-endo-conformation (bottom) the adjacent phosphates are spaced more widely apart, as compared to the 3-endo-conformation (adapted from M. Sundaralingam, in: Structure and Conformation of Nucleic Acids and Protein-Nucleic Acid Interactions, M. Sundaralingam and S.T. Rao, eds., Univ. Park Press, Baltimore, pp. 487-524, 1974).
As will be seen from what follows, exceptions from this rule always stem from a nucleic acid having an unusual structure.