![]() Go to frame view (Recommended only for
screen resolution 1024x768)
Go to frame view (Recommended only for
screen resolution 1024x768)
9.4 Reactions of Carbohydrate Moieties
If we take the reactions involving the ribose and deoxyribose of polynucleotides, two of them have been studied more or less in depth - substitution for hydrogens in hydroxyl groups and oxidation of the 2',3'-cis-glycol group in terminal nucleotides, leading to rupture of the C2'-C3' bonds. This statement should be qualified by adding that the reactions proceed in aqueous solutions. The reactions of oligo- and polynucleotides in organic solvents were studied, as a rule, in the context of development of synthetic methods. This section deals only with those reactions in organic solvents which have been studied on natural nucleic acids in search for ways to modify carbohydrate moieties in a controlled fashion.
9.4.1 Substitution for Hydrogen Atoms in Hydroxyl Groups
Acylation. Reactions of this type have been studied for RNAs having a free hydroxyl at C2, of ribose in each monomer unit in the middle of the polymer chain and an unsubstituted 2',3'-cis-glycol group at the 3' end of the molecule. DNA molecules contain only one hydroxyl at the 3' end of the polymer. Just as in RNAs, the terminal 5'-hydroxyl in DNAs usually contains a phosphate. Since the hydroxyls of carbohydrate moieties are not the only nucleophilic groups in the polynucleotide molecule, acylation may involve not only the hydroxyl groups of the sugar but also the amino groups of the heterocycle and phosphates. Therefore, selective acylation of carbohydrate moieties requires a carefully chosen set of conditions.
The optimal conditions for acetylation of the internal 2'-hydroxyls in RNAs include reaction with acetic anhydride in an aqueous solution at pH 7 (5-200 C). To ensure homogeneity of the reaction medium and better reproducibility of the results, the modification should be carried out with acetic anhydride in a 5 % aqueous solution of dimethylformamide. This procedure allows up to 70 per cent of the 2'-hydroxyls in tRNA to be easily modified without any perceptible degradation of the polymer chain.
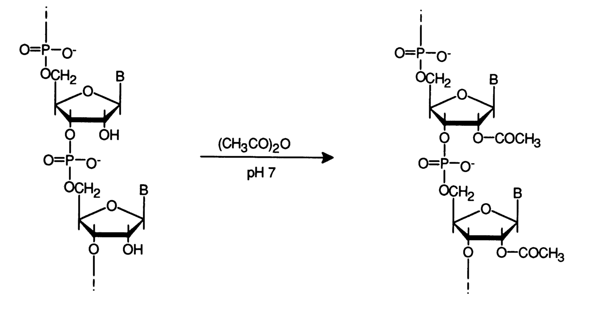
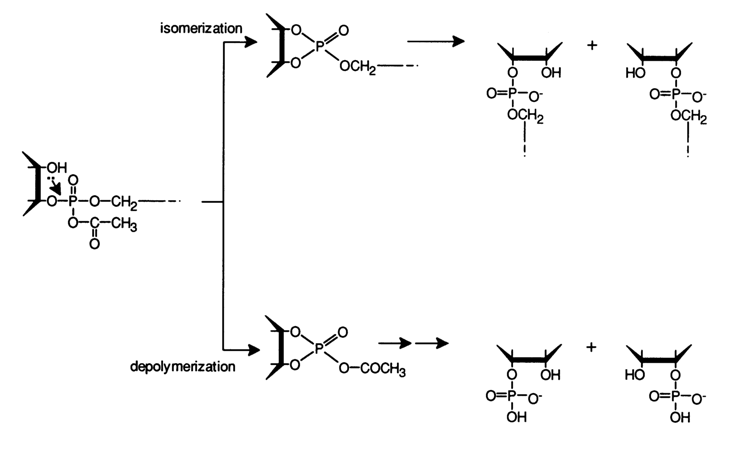
The integrity of the polynucleotide chain in tRNA during acetylation under the above conditions together with the fact that the internucleotide linkage does not undergo any isomerization indicate that the internucleotide phosphate group is not acylated. Otherwise, the following reactions would take place:
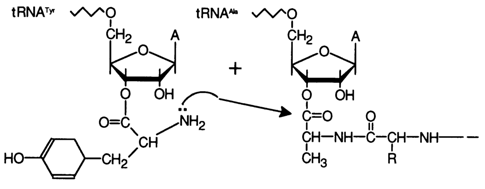
Aminoacylation. Amino acid esters of RNA and oligonucleotides have attracted a great deal of interest because tRNAs acylated with an amino acid at one of the hydroxyls of the 3'-terminal cis-glycol groups are intermediate compounds in the activation of amino acids during protein biosynthesis in ribosomes. The following scheme illustrates biosynthesis of a peptide bond in ribosomes accommodating a constantly growing polypeptide chain, linked via an ester bond to the tRNA specific for the peptide in question, as well as constantly "arriving" 3'-O-aminoacyl derivatives. As a result of growth of the peptide chain, the 3' end of the tRNA serves as the site for formation of a specific protein which has the information necessary for its synthesis stored in the mRNA; the small straight arrows show the polarity of the respective nucleic acids.

Thus, the polypeptide chain is assembled stepwise from the N-end. Studies into the properties of 3'-aminoacyl(peptidyl)-tRNA and similarly structured model compounds have shown that their ester bond is activated by the pentose moiety (such amino acid derivatives belong to the category of activated esters) and the energy it contains is quite sufficient for the peptide synthesis to take place.
At present, this process can be replicated in vitro, both with the ribosome functioning continuously on a natural or synthetic RNA template, when a polypeptide chain made up of at least several peptide bonds is formed, and in steps, with each ribosome functioning event being adequately controlled.
Along with natural participants of polypeptide synthesis on the ribosome, amino acid esters of nucleosides, nucleotides and oligonucleotides (analogues of 3'-aminoacyl-tRNA) have been successfully used as peptide acceptors in discrete events of peptide bonding, while acyloaminoacyl-oligonucleotides (analogues of peptidyl-tRNA) have been used as peptide donors. The use of such model compounds has been instrumental in elucidating the topography of the ribosomal portion (so-called peptidyl-transferase site) where the peptide bond is synthesized, and also in gleaning information on the substrate specificity of this site. In view of this and the fact that the ribosomal synthesis of peptides is widely applied to studies into the functioning of the ribosome, it would be most appropriate to develop methods for chemical synthesis of aminoacyl- and peptidyl-tRNAs and their low-molecular analogues. This is why such modification of tRNAs and oligonucleotides (as tRNA models) has received particular attention.
Selective aminoacylation of the hydroxyls in ribose becomes increasingly difficult as one goes from nucleosides to nucleotides and, to a greater degree, nucleic acids. For instance, aminoacylation of RNA may give derivatives containing an aminoacyl in the carbohydrate moiety (ester bond formed by internal 2'- or terminal 2'- or 3'-hydroxyl groups of the carbohydratephosphate backbone in RNA), in the heterocyclic base (amide bond involving the heterocyclic amino groups), and in phosphate groups (anhydride bond associated with the aminoacylation of internucleotide and terminal phosphate groups).
The aminoacylation of tRNA in organic solvents by amino acids activated at the carboxyl group yields compounds such as mixed anhydrides (reaction at phosphate groups) and amides (aminoacylation of heterocyclic bases).
In aqueous solutions, however, the aminoacylation of tRNA, just as acylation, proceeds at the hydroxyls of the carbohydrate moieties. Imidazolides of amino acids have turned out to be the best aminoacylation agents in this case. For example, condensation of N-butoxycarbonylic (Boc) aminoacylimidazolides with total tRNA from baker's yeast in a strongly aqueous medium at 200 C for several hours gives compounds with an ester (rather than anhydride or amide) type of bond. Stable and high yields were attained when the reaction was conducted in a 1 M imidazole buffer:
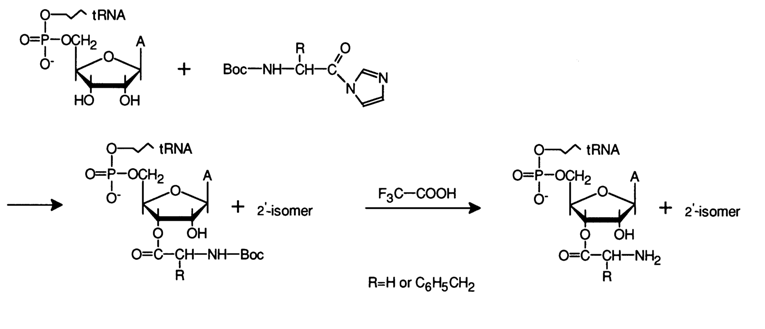
However, as the Boc group was being eliminated, the tRNA became inactivated, probably due to the rigorous conditions of the reaction. It was therefore necessary to drop this step of the synthesis. This was successfully accomplished using imidazolides of N-protonated amino acids reacting with tRNA to yield aminoacyl-tRNA in just one step:
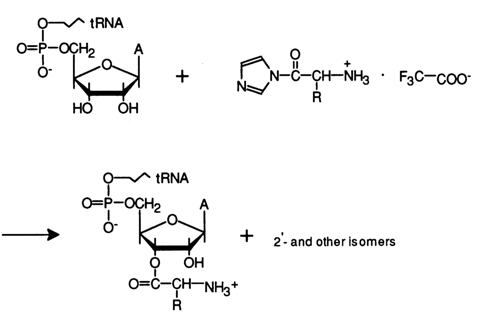
The reaction takes several hours under mild conditions (1 M aqueous buffer solution of imidazole, pH 7, 40 C). As has been demonstrated using radiometric methods, 3-10 % of the amino acid is incorporated into the terminal 3'-cis-glycol group. Moreover, the internal 2'-hydroxyl groups of ribose are aminoacylated, too.
Since special experiments with nucleotides have shown that no aminoacylation of heterocyclic bases takes place under such conditions, it may be assumed that no amide bonds are formed in the case of tRNA either. What makes this method so important is the fact that it allows the experimenter to have "hybrid" molecules with any amino acid and tRNA combination (an amino acid which is not specific with respect to a given tRNA can be added thereto).
Aminoacylation or acylation of 3'-O-aminoacyl-tRNAs (including those resulting from enzymatic phosphorylation) may yield the corresponding peptidyl(acylaminoacyl)-tRNAs. This process can be represented by the following general scheme:
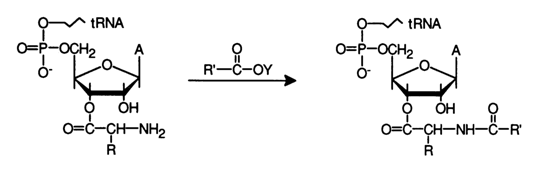
For selective acylation of amino acids use is normally made of para-nitro-phenyl
|----------------------|
(Y = C6H4NO2-p) or N-hydroxysuccinimide (Y =
NCO(CH2)2CO) esters because, as is known, such acid derivatives do
not acylate hydroxyl and amino groups in nucleic acids.
The synthesis of peptide derivatives of tRNA usually involves N-hydroxysuccinimide esters of N-methoxytritylamino acids:
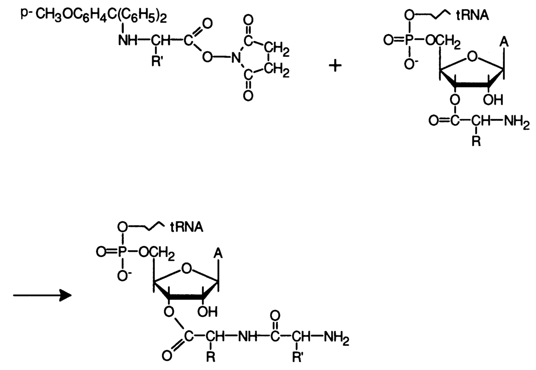
Selecting the right protective group for the amino acid is not as simple as it may seem because it should be eliminated only under very mild conditions (in order not to affect the many labile bonds in tRNA). The monomethoxytrityl group meets this criterion since it can be eliminated in the presence of 5% dichloroacetic acid at 40 C (5 min). The resulting 3'-O-peptidyl-tRNAs remain active in the ribosomal system.
The peptide chain can be built up unit by unit with dipeptide fragments being added to aminoacyl-tRNA. The yields of peptidyl-tRNA range from 80 to 90 per cent in this case.
9.4.2 Oxidation of 3'-Terminal cis-Glycol Group in RNA
The 3'-terminal cis-glycol group in RNA is readily oxidized with periodic acid. As a result, dialdehydes are formed, just as when monomer units are oxidized:
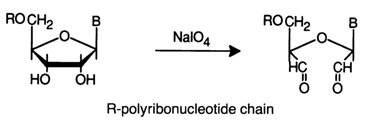
The oxidation is conducted in dilute aqueous solutions at 0-200 C. At a low temperature. the reaction proceeds quantitatively within an hour. Reduction of dialdehydes with sodium borohydride in a neutral medium yields the corresponding diols:
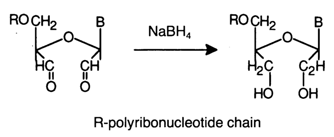
Thus, a tritium label is incorporated into RNAs or oligoribonucleotides with the aid of sodium borotritide NaBT4.
Dialdehydes easily enter into other reactions typical of carbonyl compounds. A case in point is their interaction with hydrazines and amines:
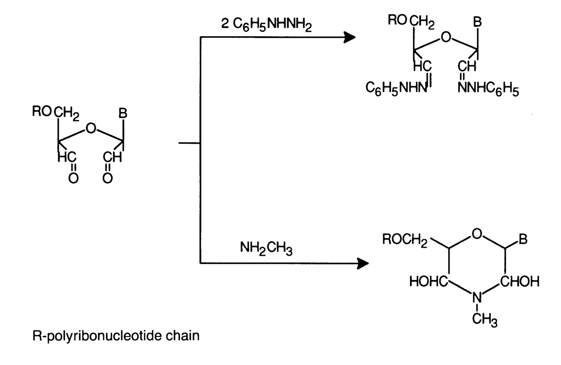
A characteristic property of periodate oxidation products is their capacity for easy degradation in an alkaline medium with
b-elimination of the phosphate group.Oxidation of the 3'-terminal glycol group with subsequent immobilization on hydrazide columns is used to isolate individual tRNAs.