![]() Go to frame view (Recommended only for
screen resolution 1024x768)
Go to frame view (Recommended only for
screen resolution 1024x768)
10.3 Ribozymes Based on Self-Cleaving RNAs
The capacity for specific self-cleavage is a rather common phenomenon among viral RNAs. As a rule, this reaction completes the replication of a viral genome, and its biological significance resides in fragmentation of an oligomeric RNA chain formed by the so-called rolling circle mechanism.
The first self-cleaving RNAs were discovered among precursors of viroids (relatively small RNAs self-replicating in plant cells) and so-called viroid satellite RNAs (emerging in plant cells only in the presence of a helper virus). The self-cleavage of multimeric forms of these RNAs occurs at a unique site and results in 2', 3'-cyclic phosphate and 5'-hydroxyl termini. Magnesium ions act as the cofactor in this highly specific hydrolytic reaction.
Comparison of the nucleotide sequences of several self-cleaving RNAs near the unique site has revealed a consensus domain accommodating the active center of the ribozyme. It consists of 50 to 60 nucleotides, and its secondary structure is represented by three helical regions joined by single-stranded segments, which is why it has been termed "hammerhead" (Fig. 10-14). The domain was found to contain 13 to 15 conserved nucleotides whose position with respect to the cleaved phosphodiester bond is strictly predetermined. The model of hammerhead secondary folding shown in Figure 10-14, constructed originally on the basis of comparative phylogenetic analysis, was subsequently supported by high-resolution NMR spectroscopy.
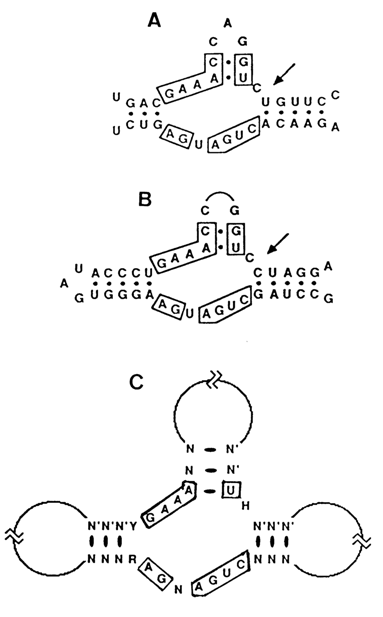
Fig. 10-14. Proposed secondary structure of hammerhead ribozymes. The arrows indicate the site of cleavage. A. Self-cleavage site of avocado sunblotch viroid (adapted from A. C. Forster and R. H. Symons, Cell, 49, 211-220 (1987). B. Self-cleavage site of the new satellite 2 RNA (adapted from L. M. Epstein and J. G. Gall, Cell, 48, 535-543 (1987). C. A hammerhead consensus structure. The boxed positions are believed to be essential due to their conservation in many hammerhead domains. Y, R, N and H designate either pyrimidine or purine and any nucleotide or any nucleotide except G, respectively (adapted from Ruffner et al., Biochemistry, 29, 1065-1072 (1990)).
It was found that the self-cleavage of such RNAs can be modeled in a rather simple system consisting of two oligoribonucleotides partially complementary with respect to each other, whose pairing reproduces the hammerhead (Fig. 10-15). These oligonucleotides had been synthesized with the aid of T7 RNA-polymerase. The cleavage of a longer oligonucleotide took place precisely where expected, and the shorter oligonucleotide 19 nucleotides long, remaining intact after the reaction was over, acted as an enzyme (ribozyme). The substrate cleavage rate reached a maximum at 550 C and remained almost the same as pH increased from 7.5 to 9.
The discovery of a simple synthetic system modeling a self-cleaving hammerhead had several important consequences. Firstly, it allowed the construction of a sufficiently simple ribozyme (Fig. 10-16) instrumental in cleavage of the internucleotide bond after C in the GCU sequence of virtually any RNA. The central portion of this ribozyme comprises an active center and, therefore, is identical in all ribozymes of this type. The specific binding of the ribozyme to a particular RNA is determined by its 3' and 5'-terminal regions. Naturally, the number and species of the base pairs formed by these regions with the RNA determines both Km and kcat of the reaction. It was then demonstrated that substitution of GUA or GUU (but not GUG) for GUC does not affect the activity of this ribozyme in any perceptible manner.
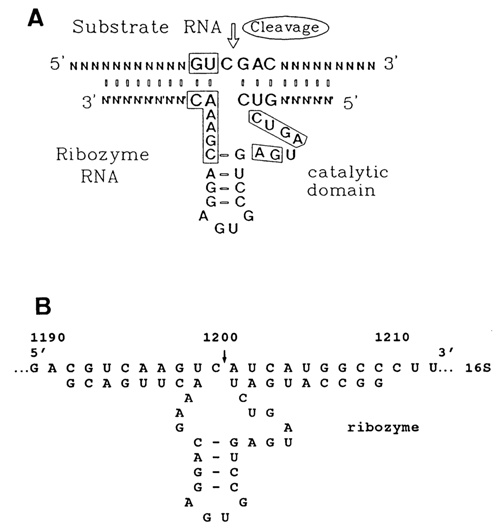
Fig. 10-15. Synthesis of a 19-nucleotide ribozyme and the structure of its complex with the substrate. Oligonucleotide O1 can be referred to as enzyme and oligonucleotide O2 as substrate only conventionally because both are involved in the hammerhead structure formation. The designations are the same as in Figure 10-14 (adapted from O. C. Uhlenbeck, Nature, 328, 596-600 (1987)).
Moreover, the ribozyme cleaved the RNA in respective complexes after CUC, AUC and UCU, albeit to a lesser degree. Therefore, such ribozymes have the potential for creating agents capable of cleaving, in a controlled manner, the genomes of RNA-containing viruses in vivo. If the problem of delivering such a ribozyme into target cells is put aside, it becomes evident that the limiting factor in this case will be the secondary and tertiary structures of the viral RNA region with which the complementary pairing of the ribozyme must take place.
Fig. 10-16. Cleavage of any exogenous RNA containing a G UC sequence by a hammerhead-like ribozyme. A. General structure of the ribozyme-substrate complex. (adapted from J. Haseloff and W L. Gerlach, Nature,334, 585-591 (1988)).
B. Cleavage of E. coli 16S ribosomal RNA (1542 nucleotide long) at single site by a ribozyme specially designed for this RNA (courtesy of S. L. Bogdanova).
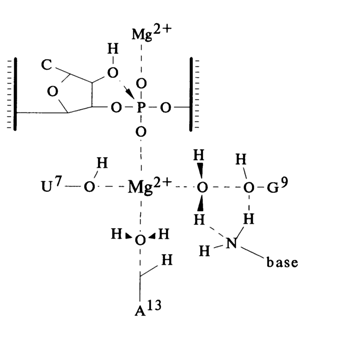
Secondly, experiments with a simplified version of the ribozyme have revealed nucleotides whose replacement or modification materially affects its activity, which was a serious step toward understanding the mechanism of self-cleavage. It was found, for example, that the hammerhead may be represented by a more rigorous consensus than expected earlier (Fig. 10-14c). It contains only 11 nucleotides whose replacement is not permissible, and all nine conserved nucleotides in the single-stranded regions of the ribozyme are necessary for self-cleavage. Furthermore, replacement of some nucleotides in the ribozyme by their deoxy analogues has shown that its activity is strongly dependent not only on the 2'-OH group next to the cleaved phosphodiester bond but also the 2'-hydroxyls of some "internal" nucleotides. At the same time, many other ribozyme residues can be substituted by the corresponding deoxynucleotides without any tangible effect on its activity. It allowed R. Cedergren and co-workers to propose the mechanism of hammerhead ribozyme action which suggests that several specific 2'-hydroxyls from both the substrate and the ribozyme coordinate a magnesium ion:
An interesting approach to identifying functionally essential ribozyme fragments is based on substitution of a phosphothioate for the usual phosphate group in its phosphodiester bonds. Among the essential features of nucleoside phosphothioates are the chirality of the phosphorus atom and the fact that the negative charge of the phosphothioate group is located primarily at the oxygen (the sulfur atom being practically neutral). T7 RNA-polymerase normally employed in the synthesis of simple ribozymes uses only the Sp isomer of NTPaS as the substrate. However, when this isomer is incorporated into the polynucleotide chain, its configuration is inverted to give the Rp isomer of the phosphothioate linkage. If the emergence of such bonds in the ribozyme alters its enzymatic activity, it may be assumed that the active center accomodates an appropriate phosphate group. Phosphothioate linkages are easily identifiable through alkylation and subsequent hydrolysis of the emerging triester group. Therefore, one of the four nucleotides in the ribozyme is statistically replaced by its phosphothio analog (under conditions when one nucleotide on the average is substituted by a molecule), then the slowly cleaving molecules are isolated for identification of the nucleotides participating in enzymatic catalysis.
All this suggests that conserved nucleotides in single-stranded regions of the hammerhead form a tertiary structure organized on principles yet to be understood. It is assumed that there appears an ensemble of functional groups retaining the bivalent metal ion near the phosphodiester bond being cleaved. As has already been mentioned, the self-cleavage of RNAs gives termini containing 2', 3'-cyclic phosphate and 5'-phosphodiester groups. Thus, the role of the metal (usually magnesium ions) seems to boil down either to enhancing the nucleophilic activity of the neighboring 2'-OH group or to making the phosphorus more electrophilic. A good analogy is provided by the specific cleavage of one of the internucleotide linkages in tRNA, catalysed by lead (Pb II) ions. As can be seen from Figure 10-17, being linked to nucleosides 59 and 60 in the TYC loop, the Pb (II) ion is a source of hydroxyl ions linked to the metal. The special system was developed on the basis of these data to isolate very simple polyribonucleotides that undergo autocatalytic cleavage with Pb2+ in the presence of Mg2+ (Fig. 10-17 B). It has also been speculated that the metal ion in the self-cleaving domain may be involved in stabilization of the transient state giving rise to 2', 3'-cyclic phosphate.
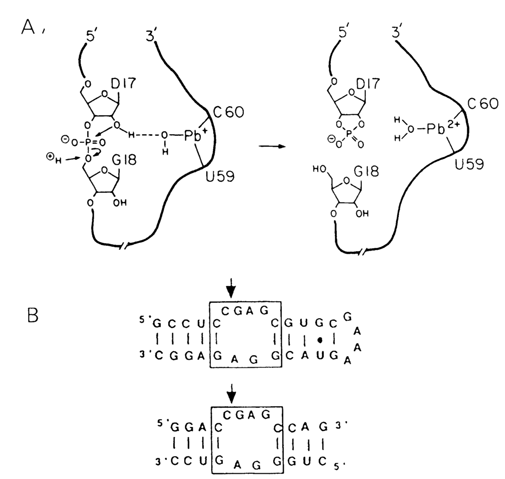
Fig. 10-17. A. Hypothetical mechanism of site-specific hydrolysis catalysed by lead ions (reproduced from T. R. Cech and B. L. Bass, Ann. Rev. Biochem., 55, 599-629 (1986)). B. Short RNA constructs that undergo Pb2+ cleavage at sites indicated by the arrows (adapted from T. Pan and 0. Uhlenbeck, Nature, 358, 560-563 (1992)).
Some other instances of self-cleavage of RNA have been reported apart from RNAs having a consensus structure of the hammerhead type. Examples are illustrated in Figure 10-18. Judging from most parameters of these reactions, they are governed basically by the same mechanism, although in each case the active center of the ribozyme has a different individual structure.
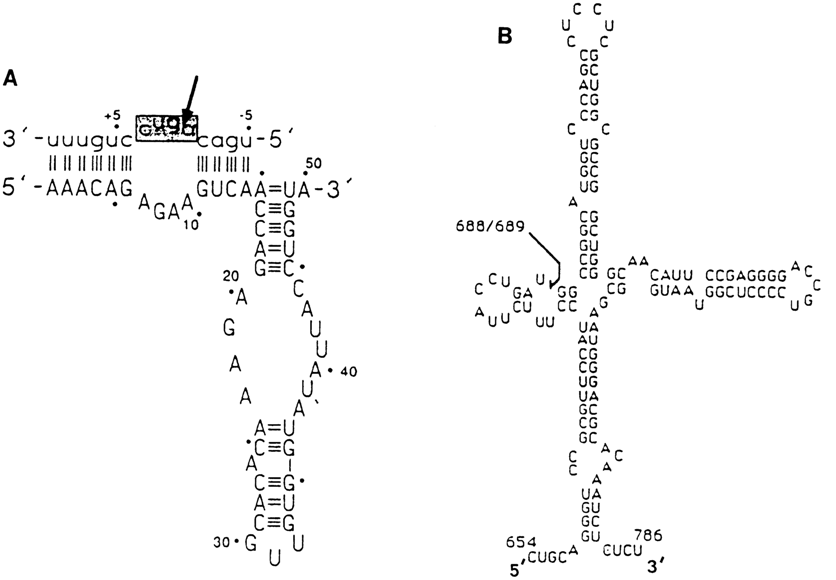
Fig. 10-18. Self-cleaving RNA domains having a secondary
structure different from the hammerhead one.
A. the hairpin ribozyme constructed on the basis of self-cleaving domain of the negative
strand of the satellite RNA of tobacco ringspot virus (adapted from B. M. Chowrira et al.,
Nature, 354, 320-322 (1991)).
B. The smallest self-cleaving fragment of human hepatitis 6 virus RNA (adapted from H.-N.
Wu et al., Proc. Natl. Acad. Sci. USA, 86, 1831-1835 (1989)). The arrows
mark cleavage sites.