![]() Go to frame view (Recommended only for
screen resolution 1024x768)
Go to frame view (Recommended only for
screen resolution 1024x768)
5.3 Nature of Intemucleotide Linkages
Studies into internucleotide linkages in polymeric nucleic acid molecules were completed successfully in the early fifties, immediately after the structure of nucleotides had been established and some properties of their derivatives (primarily esters) had been elucidated. By the same time, methods for isolating and purifying DNA and RNA had been developed so that the nature of intermonomer linkages was studied on pure, albeit markedly degraded, nucleic acid preparations.
T'he earliest information about the type of intermonomer or, as has become more common to say, internucleotide linkage came from potentiometric titration experiments. Their results indicated that RNA as well as DNA contains a single hydroxyl at each phosphate group (pKa~1). The conclusion that followed was that each nucleic acid contains a disubstituted phosphate group as a structural unit.
It was natural to assume that phosphate groups join nucleosides by crosslinks involving two of their hydroxyls, whereas one hydroxyl remains free. It only remained to find out which parts of nucleoside moieties participate in the linkage with phosphate groups.
Since nucleic acids can be deaminated in the presence of nitrous acid, it is obvious that the amino groups of pyrimidine and purine bases are not involved in internucleotide linkage. Moreover, potentiometric titration has shown that the oxo(hydroxy) groups of guanines and uracils in nucleic acids are free as well. This finding led to the conclusion that internucleotide linkages are formed by the phosphate group and the hydroxyls of carbohydrate moieties (i.e., they are essentially phosphodiester bonds), which are consequently responsible for formation of the polymer chain (nucleic acid). Thus, what is generally referred to as internucleotide linkage is, in fact, a system of bonds:
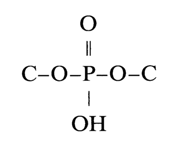
(where C is the primary or secondary carbon of the sugar). As has already been mentioned, depending on the reaction conditions, hydrolysis of DNA and RNA yields nucleotides with different positions of the phosphate group:
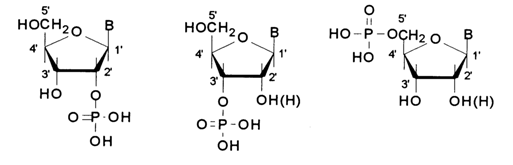
If it is assumed that all internucleotide linkages in nucleic acids are identical, then, in addition to the phosphate group, they may contain only the 3'-hydroxyl of one nucleoside moiety and the 5'-hydroxyl of the other (3'-5'bond). And if they are different, the polymer chain of DNA can accommodate three types of bonds at a time: 3'-5', 3'-3' and 5'-5'. In the case of RNA, the number of bond types must be greater because of involvement of the 2'-hydroxyl as well.
The true nature of internucleotide linkages in native DNA and RNA was established as a consequence of directed chemical and enzymatic hydrolysis of the biopolymers with subsequent isolation and identification of the resulting fragments.
5.3.1 Internucteotide Linkage in DNA
Chemical hydrolysis of DNA as a method for degrading the polymer to establish the nature of internucleotide linkage has turned out to be useless for all practical purposes. DNA is not hydrolysed at alkaline pH values, which is quite consistent with the assumption that the internucleotide linkage is phosphodiester in nature (the stability of dialkylphosphates in alkaline media has already been discussed). When treated with acid even under mild conditions, DNA is hydrolysed both at phosphodiester bonds and at N-glycosidic bonds formed by purine bases. Consequently, hydrolysis of the polymer does not yield consistent results, yet it has been possible to isolate, among products of acid DNA hydrolysis, diphosphates of pyrimidine deoxynucleosides, which are identical with synthetic deoxycytidine and deoxythymidine 3',5'-diphosphates:
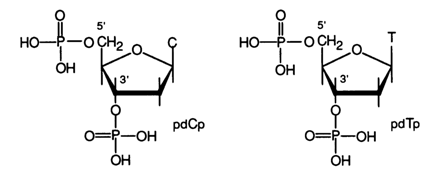
The mechanism of pdcp and pdtp formation will be discussed later; at this juncture, it is important to point out that the presence of these compounds among DNA degradation products is indicative of involvement of both hydroxyl groups, at least those of pyrimidine monomer units, in formation of the internucleotide linkage.
Enzymatic hydrolysis of DNA has turned out to be more specific. When DNA preparations were treated with snake venom phosphodiesterase (PDE), the polymer was almost completely hydrolysed to deoxynucleoside 5'-phosphates whose structure was established through comparison with the corresponding synthetic nueleotides.
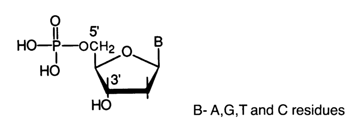
These findings suggest involvement of the 5'-hydroxyls of all four deoxynucleosides constituting DNA in internucleotide linkage. DNA in the presence of micrococcal or spleen PDE is hydrolysed in a similar manner but to 3'-phosphates.
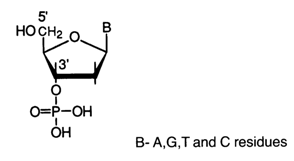
The results of DNA hydrolysis in the presence of phosphodiesterases of different specificity clearly indicate that nucleosides in DNA are linked through the medium of the phosphate group which also esterifies the hydroxyl at the secondary carbon (position 3') of one nucleoside moiety and that at the primary carbon (position 5) of another.
Thus, it has been conclusively proved that internucleotide linkage in DNA involves the phosphates group as well as 3'- and 5'-hydroxyls of the nucleosides [(a) and (b) show the directions of hydrolysis of the DNA polynucleotide chain in the presence of snake venom and spleen or micrococcal PDE, respectively]:
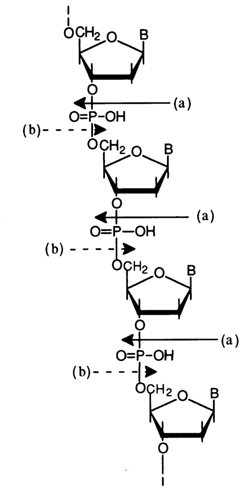
The possibility of a different polymer structure with alternating 3'-3' and 5'-5' bonds between nucleosides was ruled out because such an assumption was at variance with experimental data. For instance, such a polymer should not have been hydrolysed completely (to monomers) in the presence of snake venom PDE which selectively breaks down only nucleoside 5'-alkylphosphates. The same applies to spleen PDE which selectively hydrolyses nucleoside 3'-alkylphosphates.
5.3.2 Internucleotide Linkage in RNA
The nature of internucleotide linkage in RNA has turned out to be more challenging. Already in the earliest studies into the structure of RNA it was found to be extremely unstable during alkaline hydrolysis. The main products of alkaline hydrolysis of RNA are ribonucleoside 2'- and 3'-phosphates yielded in practically equal amounts:
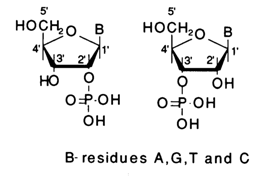
No ribonucleoside 5'-phosphates are formed in the process. These findings were hard to reconcile with the assumed phosphodiester nature of internucleotide linkage in RNA (as has already been mentioned, ordinary dialkylphosphates are relatively stable in alkaline media) and called for more comprehensive studies. An extremely important role in such studies, carried out in the early fifties by Todd and coworkers, was played by synthetic ribonucleotide alkylphosphates, which were obtained to simulate a particular type of phosphodiester bonds. The stability of such model compounds under conditions of alkaline hydrolysis has already been treated at length.
The results obtained by Todd and collaborators to elucidate the mechanisms of the transformation undergone by ribonucleotide alkylphosphates suggest that the internucleotide linkage in RNA as well as DNA involves the phosphate group and 3'- and 5'-hydroxyls of the carbohydrate moieties. A similar bond in RNA must be readily hydrolysed in an alkaline medium because the neighboring 2'-hydroxyl must catalyze this process at pH > 10, which marks the onset of ionization of the ribose hydroxyls. It should be strongly emphasized that the intermediate compounds forming during alkaline hydrolysis must be all of the four ribonucleoside 2',3'-cyclic phosphates, while the end products are the ribonucleoside 3'- and 2'-phosphates (four pairs of isomers) resulting from their hydrolysis. The following scheme illustrates alkaline hydrolysis of an RNA fragment with a random nucleotide sequence in an alkaline medium:
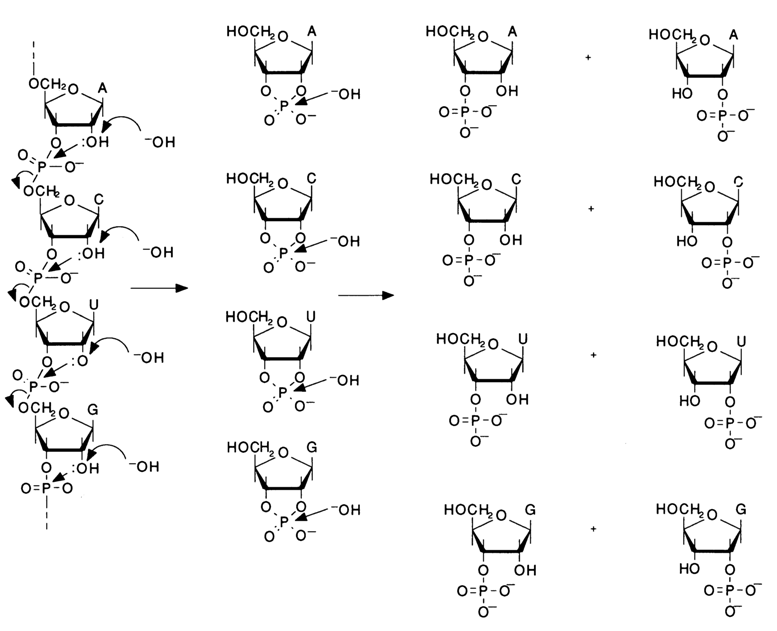
Of course, other types of internucleotide linkage could not be excluded at that stage of investigations, namely, homogeneous 2'-5' and mixed 3'-5' and 2'-3'bonds which must break in an alkaline medium to yield the same intermediates and end products. The hypothesis stating that the 3'-5' 2'-5' type of internucleotide linkage must exist in RNA was supported by two findings. When treated with an alkali, RNA is immediately hydrolysed to low-molecular fragments, which is indicative of virtually simultaneous cleavage of internucleotide linkages over the entire length of the polymer chain, rather than step by step, proceeding from the end of the chain on as cc-hydroxyls become free (such a cleavage mechanism might be expected if the monomers were linked by a 2'-3' or similar bond). Besides, mild alkaline treatment of RNA was found to inhibit the cleavage at the stage of formation of the nucleoside 2',3'-cyclic phosphates which had been isolated and identified.
The absence of dimers with 5'-5' bonding among the RNA hydrolysis products provided additional evidence in favor of the 3'-5'or 2'-5' nature of internucleotide linkage in RNA.
Thus, the results of alkaline hydrolysis showed that the number of possible internucleotide linkage types in RNA is limited but did not elucidated the structure of this polymer.
More conclusive evidence as regards the type of internucleotide linkade in RNA was provided, just as in the case of DNA, by enzymatic hydrolysis, or digestion.
RNA hydrolysis in the presence of snake venom PDE to ribonucleoside 5'-phosphates yielded a direct proof that 5'-hydroxyls participate in the phosphodiester bonding between monomer units. Later this was definitively established as a result of discovery of RNA phosphorolysis in the presence of the enzyme polynucleotide phosphorylase (PNPase), yielding ribonucleoside 5'-pyrophosphates:
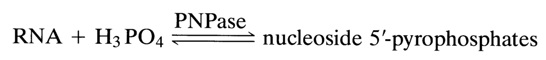
Thus, it only remained to elucidate the nature of the second hydroxyl involved in the formation of internucleotide linkage. This problem was partially solved with the aid of yet another enzyme used for controlled RNA digestion, namely, pyrimidyl ribonuclease (RNase).
As was shown earlier, this enzyme breaks down only pyrimidine ribonucleoside 3'-alkylphosphates to ribonucleoside 3'-phosphates (via the intermediate ribonucleoside 2',3'-cyclic phosphate). It was found that the enzyme acts on RNA in a similar fashion. The following example with a random nucleotide sequence illustrates how this reaction in combination with other already known ones can be used in structural studies. Experiments with any purified RNA samples have shown that the quantity of the phosphoric acid formed when the polymer is treated with pyrimidyl RNase and then with phosphomonoesterase (PME), as well as the amount of the periodic acid spent in the subsequent oxidation, is equivalent to that of the pyrimidines in the RNA sample. This suggested that at least pyrimidine nucleotides in RNA are linked with the neighboring ones only by a 3'-5' bond. This conclusion is borne out by the results of alkaline treatment of enzymatic hydrolysates of RNA after it has been exposed to RNase: in an alkaline medium, migration of the phosphate group in ribonucleoside 3'- and 2'-phosphates is impossible, and the presence of only pyrimidine ribonucleoside 3'-phosphates in the corresponding hydrolysates is a clear indication of the 3'-5' type of the internucleotide linkage in the case of pyrimidine nucleotides.
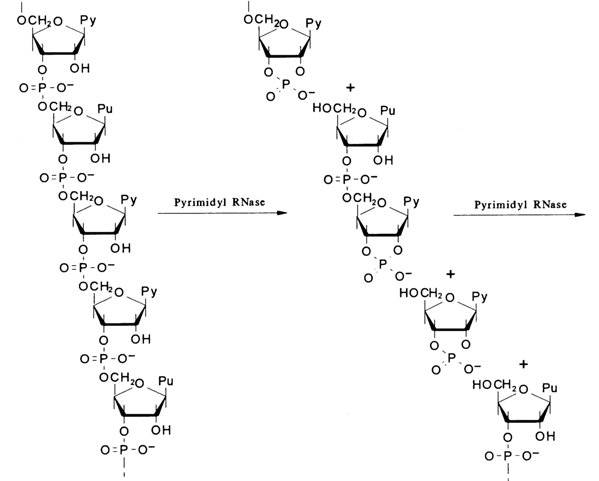
The logical assumption that the 31-5' bond must be the only intermonomer linkage in RNA was soon corroborated in experiments where spleen PDE was used to digest RNA. Just as in the case of DNA, this enzyme catalysed hydrolysis of the polynucleotide chain to nucleoside 3'-phosphates, the entire RNA being broken down to nucleotides.
It should be pointed out that guanyl (T1) RNase and also other enzymes are currently used for directed cleavage of 3'-5'bonds in RNA.
In short, it has been found that the internucleotide linkage in RNA as well as DNA is the same over the entire linear polymer chain and involves the phosphate group and 3'- and 5'-hydroxyls of the pentose. Consequently, the molecular backbone of both nucleic acids represents a carbohydrate-phosphate chain with D-ribose (RNA) or 2'-deoxy-D-ribose (DNA) and phosphate groups alternating at regular intervals. In both cases, the polymer chain contains a regularly recurring group of atoms consisting of three carbons, two oxygens and a phosphorus

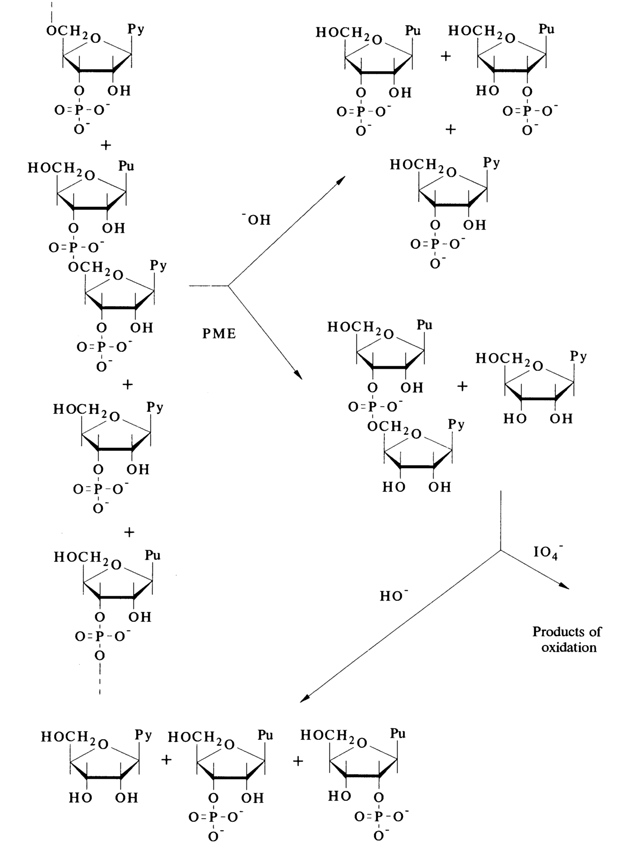
The above schematic representation of the nucleic acid polymer chain suggests that it is polar and that its terminal portions differ structurally. The terminal fragment with the free 5'-hydroxyl (or 5'-phosphate) not involved in the internucleotide linkage is known as the 5' end (or 5' terminus) of nucleic acids, while the terminal fragment with the free 3'-hydroxyl or 3'-phosphate is known as the 3'end (or 3' terminus):
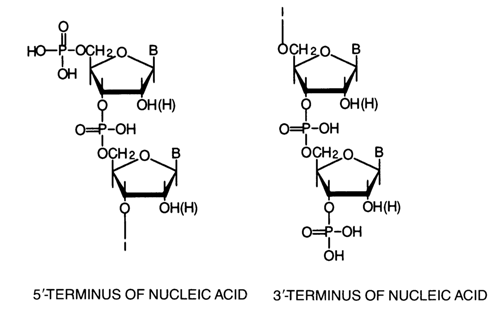
The structural formulas of DNA and RNA can be illustrated by the following scheme showing fragments of polymer chains with a random sequence of monomer units:
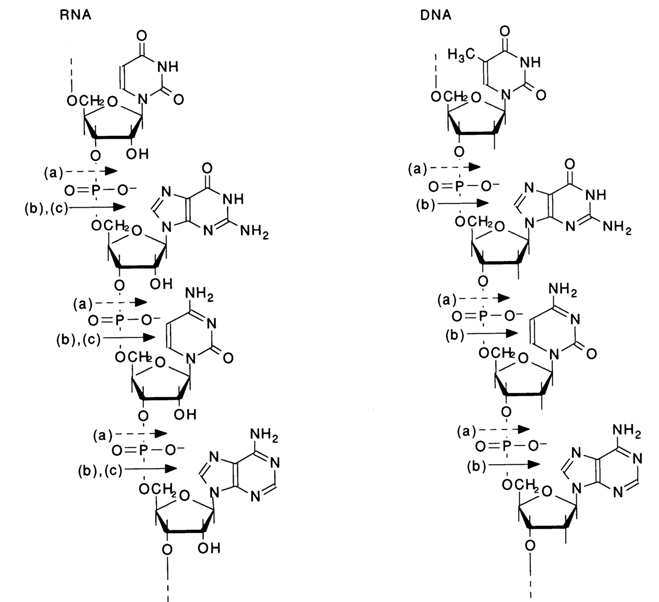
The scheme shows the sites where internucleotide linkages are cleaved in the presence of snake venom PDE [direction (a)], spleen PDE [direction (b)l and alkali [direction (c)].