![]() Go to frame view (Recommended only for
screen resolution 1024x768)
Go to frame view (Recommended only for
screen resolution 1024x768)
11.3 General Principles of Chemoenzymatic Synthesis of Duplex DNAs
As already mentioned, these principles have been formulated and implemented in the early gene syntheses by Khorana.
It became clear from the very outset that the likelihood of working out a procedure for chemical synthesis of sufficiently long DNA chains is rather small. Therefore, synthesis of extended polynucleotides had to combine chemical methods with some other approach. According to Khorana himself, this approach, which finally turned out to be so obvious and used by nature in every biological aspect of nucleic acids, of course rested on the principle of complementarity of bases.
11.3.1 Implementation of the Principle of Complementarity of Heterocyclic Bases in Nucleic Acids
Practically at the same time (in the late sixties) with development of the first effective methods for synthesizing short (approximately 10 b.p.) oligonucleotides and emergence of various complementary oligonucleotide complexes, there appeared reports announcing the discovery of the enzyme DNA ligase (polynucleotide ligase) which restores the ruptured internucleotide linkages in native DNAs. The enzyme catalyses the formation of internucleotide linkage with participation of the 5'-phosphate group and 3'-hydroxyl brought closer together by complementary interactions. This reaction can be illustrated by an example with a hypothetical DNA fragment:
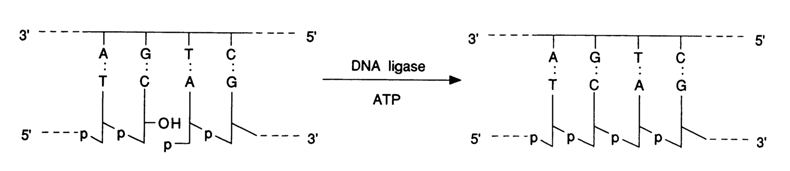
For obvious reasons DNA ligase has proved to be a real godsend for synthesists as it enabled them to combine chemically synthesized oligonucleotides within complementary complexes.
The very first attempts to use DNA ligase were aimed at finding out the minimal length of complementary oligonucleotides that can be joined together. It turned out that the latter are limited in length to 8-10 b.p., for example:
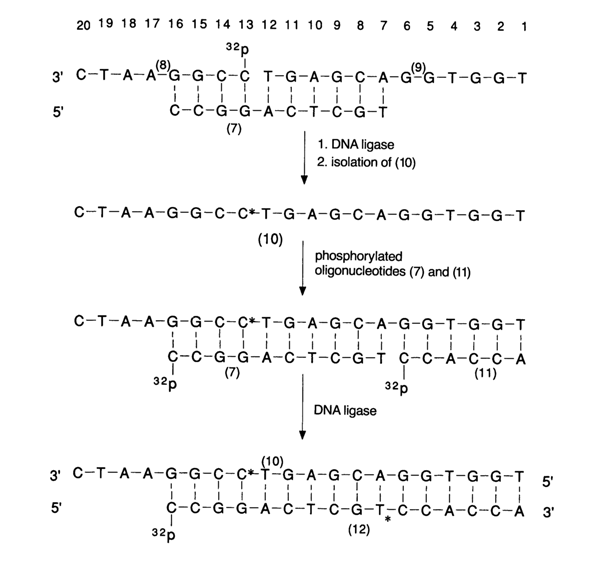
Ligation takes place when two adjacent portions of one oligonucleotide (7) are complementary to the termini of two other nucleotides (8) and (9) which, by virtue of complementary interactions, overlie the first as if it were a template. In this case, the termini are fixed in a position most favorable for covalent phosphodiester bonding.
After the ligase joins the two ends, only two polynucleotides, (7) and (10), representing a double-stranded DNA-like structure are left in the complex.
The course of the reaction is controlled by monitoring the ability of the substrate (8,) carrying the 5'-terminal phosphate group (the 5'-phosphate is usually incorporated as 32P with the aid of polynucleotide kinase) to be broken down by phosphomonoesterase to give inorganic phosphate whose quantity is measured in the reaction medium. Compounds (10) and (7) are then isolated individually to be used in subsequent buildup of the polynucleotide chain. Similarly, one can join oligonucleotide (11) to (7), in which case synthesized icosanucleotide (10) serves as the template. The ligation site and, consequently, the structure of the polynucleotides constructed in this fashion are determined (after their separation from the reaction mixture and purification) by the "closest neighbor" method.
As has already been mentioned, the starting substances for such polycondensation are oligonucleotide blocks with phosphate groups at the 5' end. Assaying by this method involves incorporation of 32P into the 5'-terminal groups [blocks (8) and (11) on the above scheme], followed by joining with the help of DNA ligase. The separated and purified polynucleotides [such as (10) and (12)] are subjected to complete hydrolysis in the presence of spleen PDE, and the nucleotide with 32P is found. The 32P-labeled phosphate group is at the 3'-terminal monomer of the oligonucleotide, to which the 5'-terminal phosphate of the second oligonucleotide has been joined with the aid of DNA ligase, and thus the ligation site can be identified. If, for example 32P becomes bound with thymidine as a result of such treatment of polynucleotides (10) and (12), the above-shown linkage of the oligonucleotide blocks (formation of the -CpT- bond) can be considered to be correct beyond any doubt.
The ligase-mediated joining does not always proceed quantitatively (sometimes its effectiveness is low) and seems to be governed by the structure of the emerging complementary complex. This is why in selecting the right conditions for enzymatic ligation of oligodeoxynucleotide blocks due regard should be given to such factors affecting the complexing process as relative concentration of the interacting components, temperature, and dilution. The effect of temperature seems to involve possible formation of a secondary structure for each substrate, for example, (7), (8) and (9), which hinders emergence of the linear DNA-like complex recognized by the enzyme.
DNA ligase may join preassembled duplexes (A, B and C) if they have sticky ends:
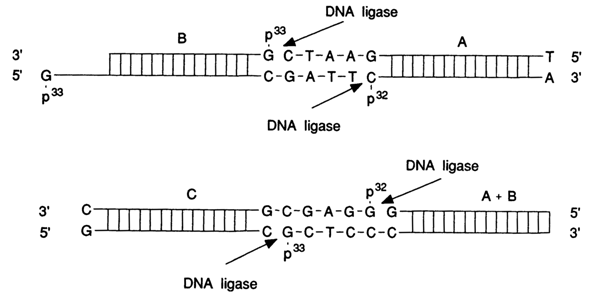
Another type of DNA ligase activity has also been discovered; namely, in
the presence of an enzyme isolated from E. coli cells infected with phage T4, two
duplexes lacking sticky ends are ligated. Such joining of blunt-ended DNA fragments
becomes more effective if RNA ligase ![]() is added to the reaction mixture.
is added to the reaction mixture.
11.3.2 Ways to Produce Duplex DNAs
It has already been pointed out that for polynucleotides to be linked together in complementary complexes (duplexes) the length of their chain must be sufficiently small (8-10 b.p.) and lie within the possibilities of chemical synthesis which existed in the sixties. Proceeding from these assumptions, Khorana formulated and then, together with his coworkers, developed a three-step procedure for duplex DNA synthesis schematically shown below:
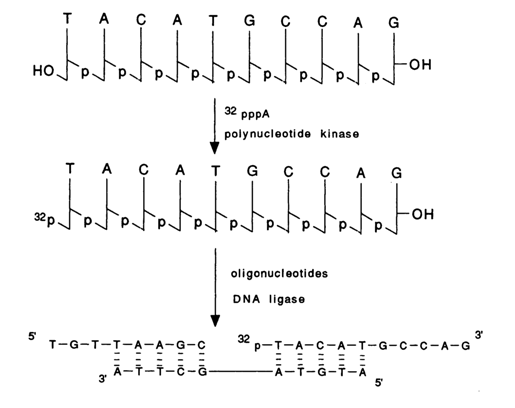
The first step is the complete chemical synthesis of oligonucleotides corresponding to both target strands. The oligonucleotides must contain free 3'- and 5'-terminal hydroxyl groups; their length usually varies from 10 to 12 nucleotides. Taken together they must represent a double-stranded DNA fragment. The overlap between fragments belonging to the opposite strands must be 5-6 b. p.
The second step involves phosphorylation of the 5'-hydroxyl groups of oligonucleotides with the aid of (
g-32P) ATP in the presence of polynucleotide kinase. Thus, all synthetic oligonucleotides become 5'-labeled compounds, which renders analysis during the third step much easier.The third step is ligation of three or more oligonucleotides in the presence of DNA ligase. To this end, segments of (7-9) and (11) types (see above scheme) with overlapping complementary sequences are mixed at an appropriate temperature and ionic strength to obtain a double-stranded complex. To ensure covalent binding of the adjacent segments resulting in a continuous duplex, the mixture is treated with DNA ligase.
In addition to the "closest neighbor" method (see above), other techniques can be used to characterize the newly formed chains by determining their length, such as polyacrylamide gel electrophoresis (PAGE). The complete nucleotide sequence of chains constructed by ligase-assisted linkage is currently determined by the Maxam-Gilbert method.
The reliability of the above-described chemoenzymatic method for producing double-stranded DNA fragments was demonstrated in Khorana's laboratory, in the synthesis of the complete 207 b. p. gene of suppressor thyrosine RNA, as well as in other laboratories, in the synthesis of protein genes and other functionally important DNA fragments.
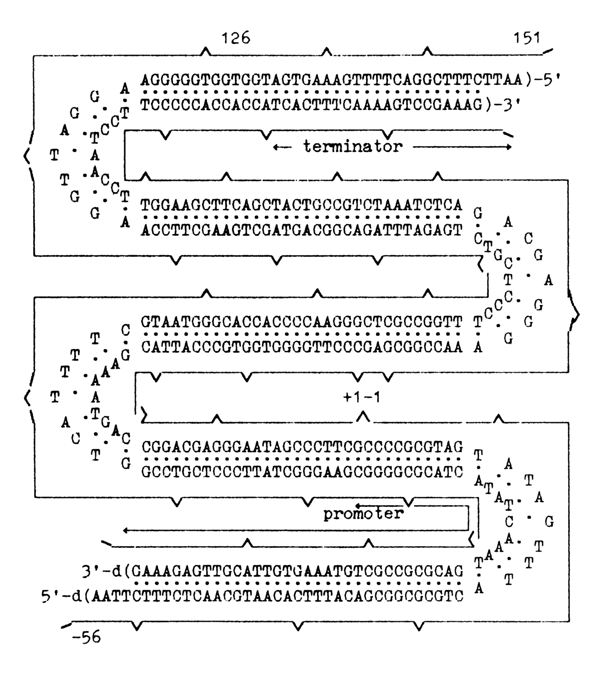
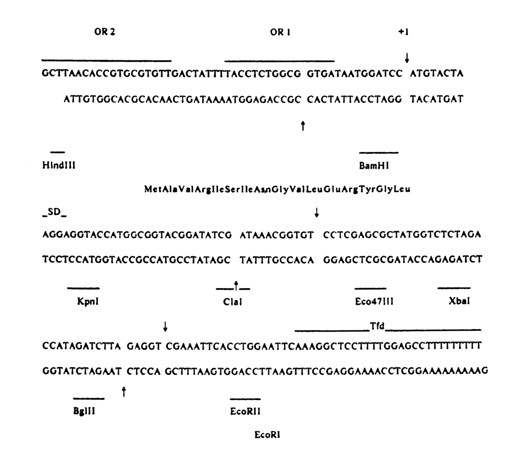
The scheme that follows shows the structure of the suppressor thyrosine tRNA gene synthesized in Khorana's laboratory, which contains, apart from the structural gene, a promoter and a terminator.
11.3.3 Enzymatic Methods for Construction of Duplex DNAs
So far, a large number of genes have been assembled from synthetic fragments to be subsequently used in microbiological synthesis of peptides and proteins by genetic engineering techniques. Table 11-3 lists such genes described in 1989.
Today, when extended (20-100 b. p) synthetic polynucleotides are available, genes are not always constructed by the method described above, often referred to as step-by-step assembly. One of the recent procedures is so-called "one-step ligation". In this case, all synthetic oligonucleotides constituting a double-stranded DNA and prepared for enzymatic ligation are placed in a vessel, T4 DNA ligase is added after heating (denaturation) and cooling, and the mixture is subjected to cloning. This is how the gene of
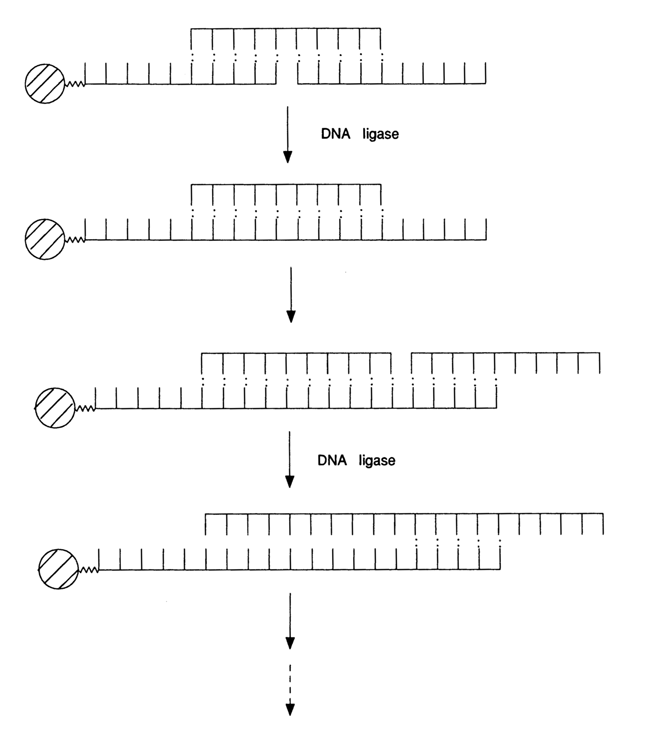
a-neoendorphin was produced in 1982, to be followed later by other genes. Sometimes, a similar approach is used, with the difference that the ligation is carried out directly in the cell in the presence of a suitable vector. Then, DNA ligase of the cell is used (Narang, 1986).Some of the reports published in recent years deal with step-by-step solid-phase construction of duplex DNAs. The basic principle behind this approach is that a terminal oligonucleotide is immobilized on an insoluble polymer with two more added to form a complementary complex, and the break in the latter is eliminated using DNA ligase. After filtration and washing, another oligonucleotide is added to the polymer and treated with DNA ligase again. Thus, both chains are built up step by step (as shown schematically below). This assembly technique is yet to be refined, but it is hard to deny the promise it holds. Once removed from the polymer (cross-hatched circle in the picture), the DNA duplex with its protruding ends is incorporated into an appropriate vector by usual means. None of the defective duplexes find their way into the vector because of lack of the right terminators. In spite of the low effectiveness of such assembly, it is quite applicable in genetic engineering. Table 11-3 shows examples of enzymatic assembly of genes from synthetic oligonucleotides using the above methods.
 11.3.4 Chemical Methods for Assembling
Duplex DNAs
11.3.4 Chemical Methods for Assembling
Duplex DNAs
Instead of an enzymatic reaction (DNA ligase) for assembling double-stranded DNAs from synthetic single-stranded oligo(poly)deoxyribonucleotides, chemical reactions resulting in internucleotide linkage can be used. This method named "chemical ligation" has been developed by Shabarova and coworkers (Moscow State University). The linkage of single-stranded fragments within a DNA duplex is achieved through activation of the phosphate groups at cleavage sites in each strand under conditions making existence of a complementary complex possible (aqueous solution, a certain ionic strength, pH, temperature). It should be emphasized here that complementary complexes are highly ordered systems in which the terminal phosphate and hydroxyl groups at cleavage sites are brought close enough to be separated by the covalent bond distance and spatially oriented in a definite way by virtue of forces stabilizing the double helix. As a consequence, one can attain chemical reaction selectivity and rates too high to be achievable for similar transformations outside the complementary complex.
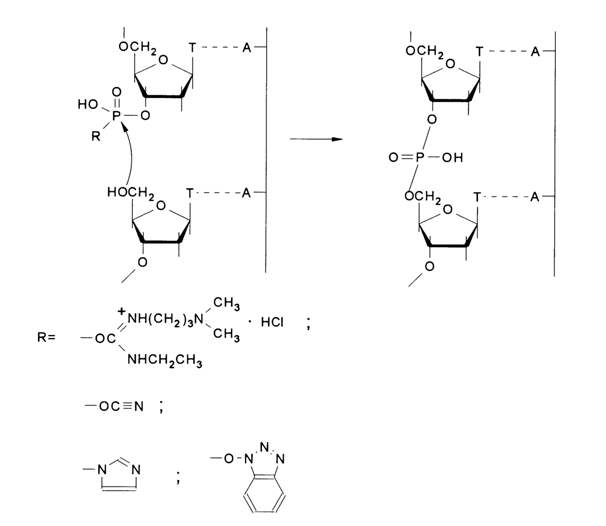
The reaction rate is increased not only because of a locally higher concentration of the reacting groups, but also as a result of their particular orientation which facilitates the transition state. As can be seen from the above scheme, two types of terminal phosphate activation are used: with the aid of condensation agents or by way of pre-activation (by obtaining potentially active oligonucleotides in advance). The first type involves reactions in which activation of the phosphorus at the junction between oligonucleotides occurs after formation of the duplex, as a result of introduction of water-soluble condensation agents, such as carbodiimides or cyanogen bromide. They easily react with the terminal monophosphate of the oligonucleotide to yield an active derivative which reacts only with the closest hydroxyl in the duplex, as is the case with enzymatic ligation. It has been established that if in the presence of carbodiimides this process takes two to three days, in the presence of cyanogen bromide its duration is as short as a minute.
By "pre-activation" (second type) is here meant use of groups covalently bound with the terminal phosphate and activating it, namely, oligonucleotide derivatives stable in aqueous solutions but exhibiting phosphorylating activity within the duplex. They include imidazolides and N-hydroxybenzotriazole phosphodiesters of oligonucleotides. Condensation in the presence of such derivatives proceeds under optimal conditions (i. e., when a stable duplex is formed and the steric and chemical factors are favorable) with good yields (up to 80-90 %). Simple methods have been elaborated for synthesis of imidazolides and other azolides of oligonucleotides as well as the more active N-hydroxybenzotriazole phosphodiesters.
Condensation agents for activating the phosphate during chemical ligation were used in studying the following chemical and steric factors of this process: position of the phosphate group in the oligonucleotide (5' and 3'), nucleophilicity of the reacting groups, position of the hydroxyl in the endo- and exo-conformation as well as its cis- and transpositions in the oxide ring of the carbohydrate moiety. It has been found that the optimal conditions for a quantitative course of chemical ligation in DNA duplexes are provided by using 3'-phosphates of natural oligonucleotides.
The above factors have been taken into consideration to produce extended DNA duplexes by chemical ligation. In the early eighties, when synthetic oligonucleotides were not as readily available, a rather successful model was proposed for DNA duplex construction, which turned out to be extremely useful not only in studying the reaction of template ligation but also in performing complete chemical synthesis of long DNA duplexes from short (9-15 b. p.) synthetic oligonucleotides. The system represents a DNA-like
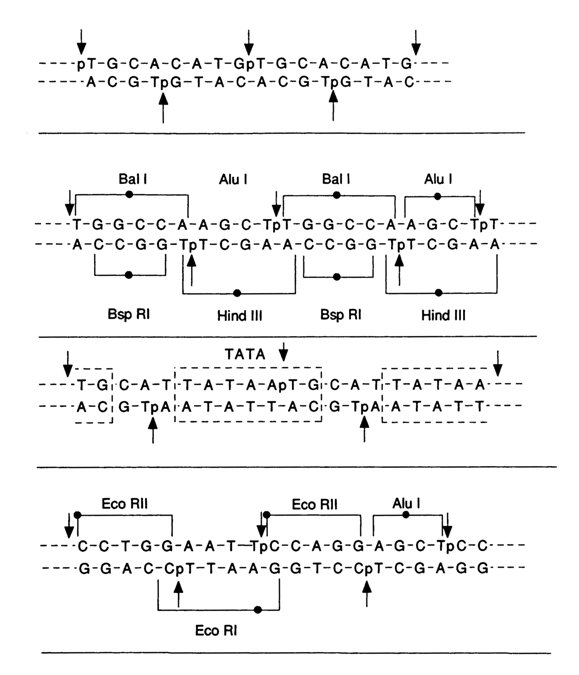
(concatemeric) complex with a recurring sequence of only one or two oligonucleotides with alternating and non-coinciding breaks in each strand. The scheme that follows illustrates such DNA duplexes. The examined complexes were different in composition and, consequently, had different theoretical stabilities at the same length of the oligomer. The physical and chemical properties of such double-stranded structures are closely similar to those of the corresponding DNAs in the B form. The above-described chemical ligation procedures were employed to assemble extended (more than 300 b. p.) duplex DNAs containing sites for recognition of various restriction endonucleases and gene promoters.
A good example of chemical ligation in double-stranded DNAs consisting of heterogeneous oligodeoxyribonucleotides (see below) is the assembly, with the aid of cyanogen bromide, of a 200 b. p. gene containing various regulatory sites and intended for modular mutagenesis while studying the functioning of different gene portions.
Arrows in the above scheme indicate the cleavage sites at which the chemical ligation takes place.
Thus, it was demonstrated that natural DNA duplexes with a predetermined primary structure can be constructed not only enzymatically using DNA ligase but also with the aid of chemical activation procedures.
An important advantage of the chemical assembly of oligonucleotides into duplexes is the possibility of introducing modified bonds (not occurring naturally) into them. As can be inferred from the references cited at the end of this chapter, when oligonucleotides modified at the 3' or 5' ends are used for ligation, phosphoamide, pyrophosphate and polyphosphate bonds formed by anomalous nucleosides (containing xylose, arabinose, lyxose) can easily and effectively be incorporated into predetermined portions of DNA, to say nothing of non-nucleoside inserts. Modified interoligomer bonds can be introduced only chemically. What makes this method especially valuable is that it is a simple and in most cases the only way to create substrates and inhibitors of nucleic exchange proteins. Now that synthetic oligonucleotides are becoming more readily available, the chemical method for constructing modified DNAs opens up wide possibilities for studying the mechanism of action of the proteins and enzymes recognizing nucleic acids. The strategy and tactics of such studies are described in the review by Ye. S. Gromova and Z. A. Shabarova published in Progress in Nucleic Acids Res. and Mol. Biol. (see references).